Introduction
A tenyésztőket és a kutatókat egyaránt érdekli a hibridizáció és az eltérő genotípusok közötti introgresszió, mint az utódok ökológiai divergenciájához vezető potenciális hajtóerő (Rieseberg et al., 2007; Arnold et al., 2012). Ezzel szemben a hibridizáció néha csökkent hibridfitnesszel, például gyengeséggel és/vagy sterilitással járhat az F1 és későbbi generációkban, még az azonos faj tagjai közötti hibridizáció esetén is.
Az F2 vagy későbbi generációkban szegregálódó csökkent hibrid életképességet és/vagy termékenységet hibrid bontásnak (HB) nevezik, amelyhez szükségszerűen recesszív allélok társulnak. Ezt a szaporodási gátat már régóta megfigyelték növényeknél és állatoknál egyaránt (Dobzhansky, 1970; Grant, 1971). E gát számos genetikai elemzéséből kiderült, hogy a gát a differenciált allélokkal rendelkező lókuszok közötti és/vagy azok közötti komplementer hatás révén valósul meg, amelyet általában Bateson-Dobzhansky-Muller (BDM) inkompatibilitásnak neveznek (Rieseberg és Willis, 2007). A HB szükségszerűen magában foglalja az intrinsic postzygotikus reprodukciós akadályokat, például a hibrid életképtelenséget (beleértve a gyengeséget, nekrózist és klorózist) és a hibrid sterilitást (a hím, nőstény vagy mindkét ivarsejtben); ezért egyes kutatók nem különböztetik meg a HB-t az F1 utódok életképtelenségétől és sterilitásától. Mindazonáltal úgy vélem, hogy a HB megfelelő osztályozás a reprodukciós akadályok számára, mivel magában foglalja a mögöttes genetikai alapjukat (azaz a recesszív allélok részvételét).
Az Arabidopsis modellnövényfajon végzett újabb vizsgálatok a HB jobb megértéséhez vezettek annak molekuláris mechanizmusait illetően (Vaid és Laitinen, 2019). Ezzel szemben, bár a rizs (Oryza faj) modellnövény, a HB genetikai alapjainak jelenlegi megértése korlátozott ebben a fajban, valószínűleg azért, mert a HB nem jelent szaporodási akadályt az F1 hibridekben, amelyekben a szülői beltenyésztett vonalakhoz képest magasabb szemtermés várható, és valószínűleg azért, mert az F2 és későbbi generációkban szegregálódó alsóbbrendű növények a fenotípus alapján könnyen kiválaszthatók a tenyészpopulációból. A recesszív öröklődés okozta genetikai térképezés nehézségei is állhatnak e korlátozás mögött.
Itt vázolom a HB-t a rizs keresztezésekben, miközben utalok az Arabidopsis vizsgálatokból származó információkra, és megvitatom, hogyan kezelik a HB-t a rizsnemesítésben.
A rizs hibridbontásának genetikai alapja
Úgy tartják, hogy a termesztett rizs, az O. sativa japonica és indica, egy fajkomplexumot alkot feltételezett ősanyjával (O. rufipogon) és vad fajokkal; azonban a köztük lévő keresztezésekben többféle szaporodási korlát, köztük a HB is megfigyelhető (Oka, 1988; Vaughan et al., 2003).
Tudomásom szerint Dr. Oka volt az első, aki leírta a HB genetikai alapjait a japonica és indica közötti intersubspecifikus keresztezésekben, amelyet két komplementer génnel magyarázott (Oka, 1957). A felelős géneket azonban nem térképezték fel a kromoszómákon, mivel akkoriban még nem álltak rendelkezésre molekuláris markerek.
A rizshibridekben a HB-t gyakran írták le intersubspecifikus (O. sativa ssp. japonica × ssp. indica) és interspecifikus (O. sativa × O. nivara és O. sativa × O. glumaepatula) keresztezésekben (Oka, 1957; Sato és Morishima, 1988; Wu et al., 1995; Li et al., 1997; Fukuoka et al., 1998, 2005; Kubo és Yoshimura, 2002, 2005; Matsubara et al., 2007a,b, 2015; Yamamoto et al., 2007; Ichitani et al., 2012;az interspecifikus keresztezésekre; Sobrizal et al., 2001; Miura et al., 2008 a fajok közötti keresztezésekre) (1. táblázat). A legtöbb ilyen HB esetet két lókuszú BDM inkompatibilitás és recesszív allélok okozták, ahol az F2 utódok 1/16-a, amely mindkét lókuszon homozigóta volt recesszív allélra, HB fenotípust mutatott; azonban a csak az egyik lókuszon heterozigóta utódok 4/16-a mutatott HB fenotípust, a keresztezési kombinációtól függően (1. táblázat és 1A. ábra). A visszakeresztezett hibridekben az egyik lókusz már rögzült a visszatérő szülő alléljaival; ezért a BCnF2 utódok 1/4-e mutatta a HB fenotípust (1. táblázat és 1B. ábra).

1. táblázat. A rizskeresztezésekben jelentett hibridbomlás genetikai alapjai.
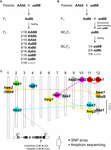
1. ábra. A hibridbontás (HB) genetikai alapja. (A) Az F2 utódokban szegregálódó HB genotípus sematikus ábrázolása. Ha a szülői genotípusok AAbb és aaBB, a domináns allél nélküli F2 utódok 1/16-a, amelyet szürke karakter jelez (azaz aabb), csökkent életképességet és/vagy termékenységet mutat, bár a többi genotípus normális. Néhány rizskeresztezésben a csak egy domináns alléllal (azaz Aabb, aaBb) rendelkező F2 utódok szintén csökkent életképességet és/vagy termékenységet mutatnak. (B) A BCnF2 utódokban szegregálódó HB genotípus sematikus ábrázolása. Továbbá a BCnF2 utódok 1/4-e csökkent életképességet és/vagy termékenységet mutat. (C) A HB alapjául szolgáló gének kromoszómális elhelyezkedése rizskeresztezésekben. A géneket az egyes vizsgálatokban végzett géntérképezés eredményei alapján nagyjából feltérképeztük. A komplementer gének egy csoportját a szaggatott vonal köti össze. A hbd1 és hwf1 komplementer géneket nem jelentették. A Yamamoto és munkatársai (2010) által közzétett példát a HB-t okozó funkcionális nukleotid-polimorfizmusra mutatjuk be, és ez az információ lehetővé teszi számunkra, hogy SNP-táblázat vagy amplikonszekvenálás segítségével felmérést végezzünk.
A rizshibridekben a HB fenotípus a keresztezési kombinációtól függően változik. Például a hbd2/hbd2 hbd3/hbd3 genotípus gyengeséget mutat, de nincs nyilvánvaló magsterilitás. A hbd4/hbd4 hbd5/hbd5 genotípus azonban gyengeséget és magsterilitást is mutat (Matsubara et al., 2007a, 2015; Yamamoto et al., 2007; 1. táblázat). Vannak olyan bonyolult esetek is, amikor gyengeséget és magsterilitást is mutató HB-t (hwe1/hwe1 hwe2/hwe2 genotípus) vagy csak magsterilitást mutató HB-t (hsa1/hsa1 hsa2/hsa2 hsa3/hsa3) találunk, ugyanannak a keresztezési kombinációnak, azaz az Asominori × IR24 keresztezésnek az ellenére (Kubo és Yoshimura, 2002, 2005; 1. táblázat). Mivel ezek a HB-k független genetikai alapokkal rendelkeznek, az F2 utódok közül legalább 5/64 mutatná a HB fenotípust ebben a keresztezésben.
A mai napig a HB-ért felelős lókuszok több csoportját térképezték fel bizonyos genomi régiókban DNS markerek segítségével a japonica és indica rizsfajták keresztezésében (Chrs 7 és 10, Fukuoka et al, 1998; Chrs 1 és 12, Wu et al., 1995; Kubo és Yoshimura, 2002; Matsubara et al., 2015; Chrs 6 és 11, Fukuoka et al., 2005; Chrs 8, 9 és 12, Kubo és Yoshimura, 2005; Chrs 2 és 11, Matsubara et al., 2007a; Yamamoto et al., 2007, 2010; Chrs 11 és 12, Ichitani et al., 2012; 1. táblázat és 1C ábra). E vizsgálatok eredményei azt mutatták, hogy a rizs HB alapjául szolgáló lókuszok egyes keresztezésekben közösek, más keresztezésekben viszont különböznek, ami a rizs genomjában való diverzifikációjukra utal. Például néhány esetben az incica fajtákkal szoros rokonságban álló O. nivara faj által hordozott HB allél egy indica fajtával osztozik ugyanazon a lókuszon; a termesztett fajtákkal szoros rokonságban álló O. glumaepatula faj által hordozott HB allél azonban nem osztozik a lókuszon egyetlen más fajtával sem (1. táblázat és 1C. ábra). Meg kell jegyezni, hogy a fent leírt HB eseteket egy sor fő hatású gén okozza; vannak azonban olyan esetek is, amelyeket egy sor kisebb hatású gén okoz, pl. enyhén csökkent vetőmagtermékenység, amelyeket a rizsnemesítésben nem lehet figyelmen kívül hagyni, és amelyeket még nem fedeztek fel.
Molecular Mechanisms Underlying Hybrid Breakdown
Autoimmun válasz: Bomblies és munkatársai (2007) számoltak be először arról, hogy az autoimmun válasz, hogy egy NB-LRR betegség-rezisztencia gén vagy R gén társul az Arabidopsis HB-hez intraspecifikus keresztezésekben, bár ez az autoimmun válasz főként nekrózis formájában fejeződik ki az F1 növényekben, és az F2 utódok HB-ja úgy tűnt, hogy a hőmérséklet által kondicionált. Arabidopsis hibridekben az autoimmun válasz által okozott HB-nek több bizonyítékát is leírták (Alcázar és mtsai., 2009, 2010). Az NB-LRR gének HB-ban való részvétele arra utal, hogy az Arabidopsis HB-hez több genomi régió is társulhat, sőt, a Chae et al. (2014) által végzett átfogó felmérés is alátámasztja ezt az elképzelést.
A rizshibridekben szintén autoimmun válasz által okozott HB-ról számoltak be egy japonica × indica keresztezésben (Yamamoto et al., 2010). Ebben az esetben a HB az F2 növényekben akkor keletkezik, amikor az indica fajta által hordozott, a kazein kináz I-et kódoló hybrid breakdown 2 (hbd2) gén kombinálódik a japonica fajta által hordozott R gének egy klaszterével. Mivel ezek a kórokozó allélok valószínűleg részben recesszív allélként viselkednek, a géntermékek is érintettek lehetnek (Matsubara et al., 2007a; Yamamoto et al., 2007, 2010).
Kettőzött gének kölcsönös elnémítása: ezt az elképzelést először elméletileg javasolták a hibrid inkompatibilitás genetikai alapjaként (Werth és Windham, 1991; Lynch és Force, 2000). A modellben az egyik populáció az egyik lókuszon funkciót veszít, a másikon pedig megtartja azt, míg a másik populáció az ellenkező hatást tapasztalja. Következésképpen az F2 zigóták 1/16-a nem rendelkezik funkcionális génekkel (Lynch és Force, 2000). A Bikard et al. (2009) és Vlad et al. (2010) által leírt HB az Arabidopsis hibridekben jó példa erre a jelenségre. Ebben a forgatókönyvben epigenetikus elnémítással járó esetekről is beszámoltak (Agorio et al., 2017; Blevins et al., 2017).
Mivel a rizsben mind a teljes genom, mind a szegmentális duplikáció tapasztalható (Wang et al., 2007; Guo et al., 2019), a reciprok elnémítás a hibrid inkompatibilitás okaként is elterjedt lehet. A mai napig azonban nincs bizonyíték a duplikált gének kölcsönös csendesítése által okozott HB-ra rizsben.
Az autoimmun választól és a kölcsönös csendesítéstől függetlenül egy DEAD-box RNS-helikáz 18 és egy másik gén (MORPHEUS MOLECULE 1 mint valószínű jelölt) közötti kölcsönhatást azonosítottak, amely HB-t okoz Arabidopsis hibridekben (Plötner et al., 2017; Vaid et al., 2020). Érdekes módon a HB fenotípus csökken az F3 és F4 generációk között, ami a génexpresszió epigenetikai szabályozására utal.
A rizshibridekben a három hibridsterilitás-a (hsa) lókusz közül Kubo és munkatársai (2016) nemrég kimutatták, hogy a hsa1 lókusz két génből áll, és hogy ezek a gének egy DUF1618 fehérjét és egy nem jellemzett, nukleotidkötő fehérjéhez némi hasonlóságot mutató fehérjét kódolnak. A fennmaradó komplementer gének, a hsa2 és hsa3 molekuláris jellemzőiről nem számoltak be.”
Hibrid törés a hagyományos rizsnemesítésben
A hagyományos rizsnemesítésben évtizedekig széles körben alkalmazták a tömeges populációs módszert. Ebben a módszerben a keresztezés után az F4 vagy F5 populációt öntermékenyítéssel ömlesztve (az egymagvú leszármazási módszer helyett), mesterséges szelekció nélkül nevelik, de a korai generációs populációt természetes és életképességi szelekciónak vetik alá (Allard, 1960; Ikehashi és Fujimaki, 1980 a részletekért). Az ömlesztett populációs módszer során várhatóan sok gyenge és/vagy steril genotípus kerül ki a populációból a fejlett generációs populáció létrehozása előtt; ezért ez a módszer korlátozott információt nyújthat a HB-ról.
A rizsnemesítők a kívánatos tulajdonságoknak a donor(ok)ból egy fajtába történő introgressziójára vagy felhalmozására gyakran végeztek visszakeresztezést vagy többszülős keresztezést, amelyet az ömlesztett populációs módszer követett. Empirikusan már régóta ismert, hogy ezek a keresztezési módszerek lehetővé teszik a szelekciós jelöltek szaporodási akadályok okozta veszteségének mérséklését, mivel ezek a módszerek gyakran csökkentik a HB-vel kapcsolatos zavaró allélkombinációk szegregációját a hibrid utódokban. A reprodukciós akadályok mérséklése ezekkel a keresztezési módszerekkel azonban elkerülhetetlenül függ a szülők HB genotípusától.
Hibrid bontás a genomikai célú nemesítésben
A referencia növénygenom szekvenciák közzététele és az újgenerációs szekvenálási technológiák fejlődése felgyorsította a molekuláris növénynemesítés fejlődését (Kole et al., 2015). Ezzel összefüggésben a növénynemesítésben fontolóra vették a genomika-asszisztált nemesítést, például a genom-szerte használt DNS-markerek genotípusain alapuló genomikai szelekciót (Spindel és Iwata, 2018).
A saját beporzású növények genomika-asszisztált nemesítésében a fejlett generációs populációkat, például a rekombináns beltenyésztett vonalakat gyakran használják referenciapopulációként, amelyekből genom-szerte használt genotípus- és fenotípusadatokat nyernek. Ezt követően a korai generációk utódain (pl. F2) kizárólag a markergenotípusokon alapuló szelekciót végeznek. Ebben a szelekciós sémában meg kell jegyezni, hogy a referenciapopuláció általában nem szolgáltat információt a HB genomi régiójáról, mivel a gyenge és/vagy steril utódokat a keresztezés után a korai generációkban ki kellett volna választani. Ezért, ha ilyen szelekciós sémát alkalmazunk, néhány fontos szelekciós jelöltről le kell mondanunk. Alternatív megoldásként nemkívánatos jelölteket szelektálhatunk úgy, hogy a HB a későbbi generációkban válik nyilvánvalóvá.
Elbeszélgetés
A témával kapcsolatos korlátozott számú vizsgálat ellenére úgy tűnik, hogy a rizskeresztezésekben a HB alapjául szolgáló lókuszok inkább diverzifikáltak, mint közösek (1. táblázat és 1. ábra). A fent leírtak szerint a HB-hez kapcsolódó allélok keresztszülők közötti eloszlásáról szóló információnak előfeltételnek kell lennie a rizsnemesítésben, különösen a genomikai támogatású nemesítés hatékonyságának maximalizálásához. Ezért szükség van a HB további kimutatására rizskeresztezésekben és a felelős gének feltérképezésére a rizs genomjában. Arabidopsis hibridekben az R génekre és a velük kölcsönhatásban lévő génekre vonatkozó ilyen adatokat széles körben felmérték (Alcázar et al., 2010, 2014; Chae et al., 2014). A Yamamoto és munkatársai (2010) által egy rizshibridben a hbd2 génről készített jelentés jó példa az SNP-táblázatok vagy amplikonszekvenálás segítségével végzett átfogó felmérésre (1C. ábra). A hbd3 és a hsa1 gének szintén jelöltek az ilyen típusú felmérésre. Végül olyan kritériumok kidolgozása, amelyek lehetővé teszik a HB előrejelzését a szülői vonalak genomikai információi alapján, fontos eszközként fog szolgálni a genomika-alapú nemesítéshez. Még ha az okozó tényezőt nem is sikerült egyetlen génként azonosítani, a szorosan összefüggő markerek, például az egynukleotid-polimorfizmusok, hatékonyak lesznek a HB-val összefüggő haplotípusok osztályozásában, mivel a termesztett rizs kapcsolati egyenlőtlensége a becslések szerint 100-200 kb-ig terjed, bár a vadon élő fajok (például az O. rufipogon) több tíz kb-ra is kiterjedhet (Mather et al., 2007; McNally et al., 2009; Huang et al., 2010). Ez az információ lehetővé teszi a hatékonyabb és eredményesebb keresztezés-kombináció tervezését.
A HB-hez kapcsolódó lókuszok feltérképezéséhez, valamint a hagyományos kapcsoltsági térképezéshez (például a mennyiségi tulajdonságlókuszok elemzéséhez) hasznos lehet a következő generációs szekvenálással végzett bolyhos szegregátumelemzés, mivel a HB szegregátumok gyakran megkülönböztethető tulajdonságokat mutatnak (olyan tulajdonságok tekintetében, mint a növénymagasság, a tőrök száma és a termékenység) a normál növekedésű szegregátumoktól az egyes keresztezési populációkban. A laboratóriumi és rizsföldön végzett fenotipizálásban fontos szerepet játszhat a digitális kamerával és drónrakodással történő képalkotó adatgyűjtés is. Ezek az erőfeszítések nemcsak a rizsnemesítők, hanem az evolúcióbiológusok számára is értékes információkkal fognak szolgálni.
Ezeken túlmenően, ha egy HB-asszociált allélt azonosítanak, a génbontás ionnyalábbal vagy genomszerkesztéssel segíthet a HB leküzdésében, amint azt a rizs F1 hibridsterilitása példázza (Koide és mtsai., 2018; Xie és mtsai, 2019).
Adatok elérhetőségi nyilatkozata
Az ebben a tanulmányban bemutatott összes adatkészletet a cikk/kiegészítő anyag tartalmazza.
A szerző hozzájárulása
A szerző megerősíti, hogy ő a munka egyedüli szerzője, és jóváhagyta annak közzétételét.
Érdekellentét
A szerzők kijelentik, hogy a kutatást olyan kereskedelmi vagy pénzügyi kapcsolatok hiányában végezték, amelyek potenciális összeférhetetlenségként értelmezhetők.
Köszönet
Ezt a tanulmányt Dr. Darshan Brar emlékének ajánlom. Ő egy kiváló rizsnemesítő volt, aki erőteljesen vitte be a vadon élő fajok által hordozott hasznos géneket a fajtákba a reprodukciós korlátokon túl.
Allard, R. W. (1960). A növénynemesítés alapelvei. New York, NY: John Wily and Sons, Inc.
Google Scholar
Dobzhansky, T. (1970). Az evolúciós folyamat genetikája. New York, NY: Columbia University Press.
Google Scholar
Grant, V. (1971). Plant Speciation. New York, NY: Columbia University Press.
Google Scholar
Ikehashi, H., and Fujimaki, H. (1980). “Modified bulk population method for rice breeding,” in Proceedings of the International Rice Research Conference on Innovative approaches to rice breeding, (Los Baños, PA: International Rice Research Institute), 163-182.
Google Scholar
Oka, H.-I. (1988). A termesztett rizs eredete. Tokyo: JSSP.
Google Scholar