Evaluación |Biopsicología |Comparativa |Cognitiva |Desarrollo |Lenguaje |Diferencias individuales |Personalidad |Filosofía |Social |
Métodos |Estadística |Clínica |Educativa |Industrial |Profesional |Psicología mundial |
Biológica:Genética del comportamiento – Psicología evolutiva – Neuroanatomía – Neuroquímica – Neuroendocrinología – Neurociencia – Psiconeuroinmunología – Psicología fisiológica – Psicofarmacología(Índice, Esquema)
| Cerebro: Hipotálamo | ||
|---|---|---|
 |
||
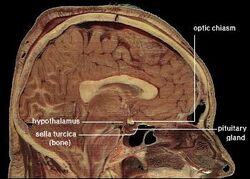
| Localización del hipotálamo humano | ||
 |
||
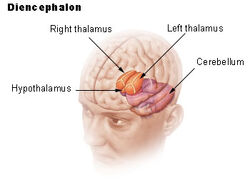
| Diencéfalo | ||
| Latino | hipotálamo | |
| Gray’s | sujeto #189 812 | |
| Parte de | ||
| Componentes | ||
| Arteria | ||
| Vein | ||
| hier-358 | ||
| MeSH | A08.186.211.730.385.357 | |
El hipotálamo une el sistema nervioso con el sistema endocrino a través de la glándula pituitaria (hipófisis). El hipotálamo, (del griego ὑποθαλαμος = bajo el tálamo) está situado debajo del tálamo, justo encima del tronco cerebral. Esta glándula ocupa la mayor parte de la región ventral del diencéfalo. Se encuentra en el cerebro de todos los mamíferos. En los seres humanos, tiene aproximadamente el tamaño de una almendra.
El hipotálamo regula ciertos procesos metabólicos y otras actividades del sistema nervioso autónomo. Sintetiza y secreta neurohormonas, a menudo denominadas hormonas liberadoras del hipotálamo, y éstas, a su vez, estimulan o inhiben la secreción de las hormonas hipofisarias.
El hipotálamo controla la temperatura corporal, el hambre, la sed, la fatiga, la ira y los ciclos circadianos.
- Entradas
- Estímulos olfativos
- Estímulos transmitidos por la sangre
- Esteroides
- Entradas neuronales
- Núcleos
- Salidas
- Proyecciones neurales
- Hormonas endocrinas
- Control de la ingesta de alimentos
- Dimorfismo sexual
- Respuestas a los esteroides ováricos
- Esteroides gonadales en la vida neonatal de las ratas
- Los andrógenos en los primates
- Otras influencias sobre el desarrollo hipotalámico
- Efectos del envejecimiento en el hipotálamo
- Véase también
- Imágenes adicionales
- Referencias & Bibliografía
- Papeles
- Material adicional
- Papeles
Entradas
El hipotálamo es una región muy compleja en el cerebro de los seres humanos, e incluso pequeños núcleos dentro del hipotálamo están involucrados en muchas funciones diferentes. El núcleo paraventricular, por ejemplo, contiene neuronas de oxitocina y vasopresina (también llamada hormona antidiurética) que se proyectan a la hipófisis posterior, pero también contiene neuronas que regulan la secreción de ACTH y TSH (que se proyectan a la hipófisis anterior), los reflejos gástricos, el comportamiento maternal, la presión sanguínea, la alimentación, las respuestas inmunitarias y la temperatura.
El hipotálamo coordina muchos ritmos circadianos hormonales y de comportamiento, patrones complejos de salidas neuroendocrinas, mecanismos homeostáticos complejos y muchos comportamientos importantes. Por lo tanto, el hipotálamo debe responder a muchas señales diferentes, algunas de las cuales se generan externamente y otras internamente. Por lo tanto, está ricamente conectado con muchas partes del SNC, incluyendo la formación reticular del tronco cerebral y las zonas autonómicas, el cerebro anterior límbico (particularmente la amígdala, el tabique, la banda diagonal de Broca y los bulbos olfatorios, y la corteza cerebral).
El hipotálamo responde a:
- La luz: la duración del día y el fotoperiodo para regular los ritmos circadianos y estacionales
- Estímulos olfativos, incluidas las feromonas
- Esteroides, incluidos los esteroides gonadales y los corticoesteroides
- Información transmitida por vía neural que procede, en particular, del corazón, el estómago y el tracto reproductivo
- Ingresos autonómicos
- Estímulos transmitidos por la sangre, incluyendo la leptina, la grelina, la angiotensina, la insulina, las hormonas hipofisarias, las citoquinas, las concentraciones plasmáticas de glucosa y la osmolaridad, etc
- El estrés
- Invita a los microorganismos a aumentar la temperatura corporal, reajustando el termostato del cuerpo hacia arriba.
Estímulos olfativos
Los estímulos olfativos son importantes para la reproducción y la función neuroendocrina en muchas especies. Por ejemplo, si se expone a un ratón preñado a la orina de un macho «extraño» durante un período crítico después del coito, el embarazo fracasa (efecto Bruce). Así, durante el coito, una hembra de ratón forma una «memoria olfativa» precisa de su pareja que persiste durante varios días.Las señales feromonales ayudan a la sincronización del celo en muchas especies; en las mujeres, la sincronización de la menstruación también puede surgir de las señales feromonales, aunque algunos dudan del papel de las feromonas en los humanos.
Estímulos transmitidos por la sangre
Las hormonas peptídicas ejercen una importante influencia sobre el hipotálamo, y para ello deben sortear la barrera hematoencefálica. El hipotálamo está delimitado en parte por regiones cerebrales especializadas que carecen de una barrera hematoencefálica eficaz; el endotelio capilar de estos lugares está fenestrado para permitir el paso libre incluso de grandes proteínas y otras moléculas. Algunos de estos lugares son los sitios de neurosecreción: la neurohipófisis y la eminencia media. Sin embargo, otros son lugares en los que el cerebro toma muestras de la composición de la sangre. Dos de estos lugares, el órgano subfornical y el OVLT (órgano vascular de la lámina terminal) son los llamados órganos circunventriculares, donde las neuronas están en íntimo contacto con la sangre y el LCR. Estas estructuras están densamente vascularizadas y contienen neuronas osmorreceptoras y receptoras de sodio que controlan la bebida, la liberación de vasopresina, la excreción de sodio y el apetito de sodio. También contienen neuronas con receptores para la angiotensina, el factor natriurético auricular, la endotelina y la relaxina, cada uno de los cuales es importante en la regulación del equilibrio de líquidos y electrolitos. Las neuronas del OVLT y del SFO se proyectan al núcleo supraóptico y al núcleo paraventricular, y también a las áreas hipotalámicas preópticas. Los órganos circunventriculares también pueden ser el lugar de acción de las interleucinas para provocar tanto la fiebre como la secreción de ACTH, a través de los efectos sobre las neuronas paraventriculares.
No está claro cómo todos los péptidos que influyen en la actividad hipotalámica obtienen el acceso necesario. En el caso de la prolactina y la leptina, existen pruebas de una captación activa en el plexo coroideo desde la sangre al LCR. Algunas hormonas hipofisarias ejercen una influencia de retroalimentación negativa sobre la secreción hipotalámica; por ejemplo, la hormona del crecimiento retroalimenta el hipotálamo, pero no está claro cómo entra en el cerebro. También hay pruebas de la acción central de la prolactina y la TSH.
Esteroides
El hipotálamo contiene neuronas sensibles a los esteroides gonadales y a los glucocorticoides – (las hormonas esteroides de la glándula suprarrenal, liberadas en respuesta a la ACTH). También contiene neuronas especializadas sensibles a la glucosa (en el núcleo arqueado y el hipotálamo ventromedial), que son importantes para el apetito. El área preóptica contiene neuronas termosensibles; éstas son importantes para la secreción de TRH.
Entradas neuronales
El hipotálamo recibe muchas entradas del tronco encefálico; en particular, del núcleo del tracto solitario, el locus coeruleus y la médula ventrolateral. La secreción de oxitocina en respuesta a la lactancia o a la estimulación vagino-cervical está mediada por algunas de estas vías; la secreción de vasopresina en respuesta a los estímulos cardiovasculares procedentes de los quimiorreceptores del seno carotídeo y del arco aórtico, y de los receptores de volumen auricular de baja presión, está mediada por otras. En la rata, la estimulación de la vagina también provoca la secreción de prolactina, lo que da lugar a un pseudoembarazo tras un apareamiento infértil. En la coneja, el coito provoca una ovulación refleja. En la oveja, la estimulación cervical en presencia de altos niveles de estrógeno puede inducir un comportamiento maternal en una oveja virgen. Todos estos efectos están mediados por el hipotálamo, y la información se transmite principalmente por vías espinales que se transmiten en el tronco cerebral. La estimulación de los pezones estimula la liberación de oxitocina y prolactina y suprime la liberación de LH y FSH.
Los estímulos cardiovasculares son transportados por el nervio vago, pero éste también transmite una variedad de información visceral, incluyendo, por ejemplo, las señales derivadas de la distensión gástrica para suprimir la alimentación. Una vez más, esta información llega al hipotálamo a través de relés en el tronco cerebral.
Núcleos
Los núcleos hipotalámicos incluyen los siguientes:
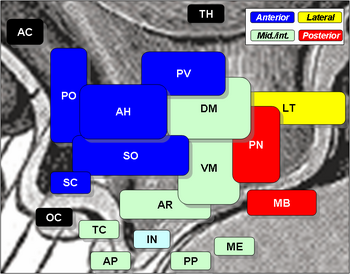
Núcleos hipotalámicos
| Región | Área | Núcleo | Función |
| Anterior | Medial | Núcleo preóptico medial |
|
| Núcleo supraóptico núcleo supraóptico (SO) |
|
||
| núcleo paraventricular (PV) |
|
||
| Núcleo hipotalámico anterior (AH) |
|
||
| Núcleo supraquiasmático (SC) |
|
||
| Núcleo preóptico lateral | Núcleo preóptico lateral | ||
| Núcleo lateral (LT) |
|
||
| Parte del núcleo supraóptico (SO) |
|
||
| Tuberal | Medial | Núcleo hipotalámico dorsomedial (DM) |
|
| Núcleo ventromedial (VM) |
|
||
Núcleo arcuato (AR)
|
|||
| Lateral | Núcleo lateral (LT) |
|
|
| Núcleos tubulares laterales | |||
| Posterior | Medial | Núcleos mamarios (parte de los cuerpos mamarios) (MB) |
|
| Núcleo posterior (NP) |
|
||
| Núcleo lateral (LT) |
Ver también: Núcleo preóptico ventrolateral
Salidas
Las salidas del hipotálamo pueden dividirse en dos categorías: proyecciones neurales, y hormonas endocrinas.
Proyecciones neurales
La mayoría de los sistemas de fibras del hipotálamo discurren en dos direcciones (bidireccionales).
- Las proyecciones a las áreas caudales al hipotálamo pasan por el haz medial del cerebro anterior, el tracto mammillotegmental y el fascículo longitudinal dorsal.
- Las proyecciones a las áreas rostrales al hipotálamo son llevadas por el tracto mamilotalámico, el fórnix y la estría terminal.
Hormonas endocrinas
El Hipotálamo afecta al sistema endocrino y gobierna el comportamiento emocional, como, la ira y la actividad sexual. La mayoría de las hormonas hipotalámicas generadas se distribuyen a la hipófisis a través del sistema portal hipofisario. El hipotálamo mantiene la homeostasis, lo que incluye la regulación de la presión arterial, la frecuencia cardíaca y la temperatura.
Las principales hormonas hipotalámicas son:
| Nombre | Otros nombres | Abreviaturas | Localización | Función |
|---|---|---|---|---|
| Factor liberador de corticotropina, Corticoliberina | CRH, CRF | neuronas neuroendocrinas parvocelulares del núcleo paraventricular | con vasopresina, estimula la pituitaria anterior para secretar ACTH | |
| Dopamina | Hormona inhibidora de la prolactina | DA, PIH | neuronas neuroendocrinas del núcleo arcuato | inhibe la secreción de prolactina de la pituitaria anterior |
| Hormona liberadora de gonadotropina | Hormona liberadora de hormona luteinizante | GnRH, LHRH | Neuronas neuroendocrinas en los núcleos preóptico medial y arqueado | Estimula la pituitaria anterior para que segregue LH y FSH |
| Hormona liberadora de la hormona del crecimiento | Factor liberador de la hormona del crecimiento, somatocrinina | GHRH, GHRF, GRF | Neuronas neuroendocrinas del núcleo arcuato | Estimula la pituitaria anterior para que segregue la hormona del crecimiento |
| Melatonina | ||||
| Somatostatina | Hormona inhibidora del crecimiento.hormona inhibidora del crecimiento, Factor inhibidor de la liberación de somatotropina | SS, GHIH, SRIF | neuronas neuroendocrinas del núcleo periventricular | inhibe la secreción de la hormona del crecimiento de la hipófisis anterior |
| Hormona liberadora de tirotropina | Factor liberador de tirotropina, Tiroliberina, Protirelina | TRH, TRF | neuronas neuroendocrinas parvocelulares en los núcleos hipotalámicos paraventricular y anterior | estimula la pituitaria anterior para que secrete TSH |
Ver también: Hipocretina
Control de la ingesta de alimentos
La parte lateral extrema del núcleo ventromedial del hipotálamo es responsable del control de la ingesta de alimentos. La estimulación de esta zona provoca un aumento de la ingesta de alimentos. La lesión bilateral de esta zona provoca el cese total de la ingesta de alimentos. La parte medial del núcleo tiene un efecto de control sobre la parte lateral. La lesión bilateral de la parte medial del núcleo ventromedial provoca hiperfagia y obesidad en el animal. La lesión posterior de la parte lateral del núcleo ventromedial en el mismo animal produce el cese completo de la ingesta de alimentos.
Existen diferentes hipótesis relacionadas con esta regulación:
- Hipótesis lipostática: esta hipótesis sostiene que el tejido adiposo produce una señal humoral que es proporcional a la cantidad de grasa y actúa sobre el hipotálamo para disminuir la ingesta de alimentos y aumentar la producción de energía. Se ha evidenciado que una hormona, la leptina, actúa sobre el hipotálamo para disminuir la ingesta de alimentos y aumentar el gasto energético.
- Hipótesis de los gutpéptidos – se afirma que las hormonas gastrointestinales como la Grp, los glucagones, la CCK y otras inhiben la ingesta de alimentos. El alimento que entra en el tracto gastrointestinal desencadena la liberación de estas hormonas que actúan en el cerebro para producir saciedad. El cerebro contiene receptores CCK-A y CCK-B.
- Hipótesis glucostática – la actividad del centro de saciedad en los núcleos ventromediales se rige probablemente por la utilización de la glucosa en las neuronas. Se ha postulado que cuando su utilización de la glucosa es baja y, en consecuencia, cuando la diferencia de glucosa en sangre arteriovenosa a través de ellas es baja, la actividad a través de las neuronas disminuye. En estas condiciones, la actividad del centro de alimentación no se controla y el individuo siente hambre. La ingesta de alimentos aumenta rápidamente mediante la administración intraventricular de 2-deoxiglucosa, por lo que disminuye la utilización de la glucosa en las células.
- Hipótesis termoestática – según esta hipótesis, una disminución de la temperatura corporal por debajo de un punto de ajuste determinado estimula el apetito, mientras que un aumento por encima del punto de ajuste lo inhibe.
Dimorfismo sexual
Varios núcleos hipotalámicos son sexualmente dimórficos, es decir, existen claras diferencias tanto en su estructura como en su función entre machos y hembras.
Algunas diferencias son evidentes incluso en la neuroanatomía macroscópica: la más notable es el núcleo sexualmente dimórfico dentro del área preóptica, que sólo está presente en los machos. Sin embargo, la mayoría de las diferencias son cambios sutiles en la conectividad y la sensibilidad química de determinados conjuntos de neuronas.
La importancia de estos cambios puede reconocerse por las diferencias funcionales entre machos y hembras. Por ejemplo, el patrón de secreción de la hormona del crecimiento es dimórfico desde el punto de vista sexual, y ésta es una de las razones por las que, en muchas especies, los machos adultos son mucho más grandes que las hembras.
Respuestas a los esteroides ováricos
Otros dimorfismos funcionales llamativos se dan en las respuestas conductuales a los esteroides ováricos del adulto. Los machos y las hembras responden de forma diferente a los esteroides ováricos, en parte porque la expresión de las neuronas sensibles a los estrógenos en el hipotálamo es sexualmente dimórfica, es decir, los receptores de estrógenos se expresan en diferentes conjuntos de neuronas.
Los estrógenos y la progesterona pueden influir en la expresión de genes en neuronas concretas o inducir cambios en el potencial de la membrana celular y en la activación de quinasas, lo que conduce a diversas funciones celulares no genómicas. Los estrógenos y la progesterona se unen a sus receptores hormonales nucleares correspondientes, que se trasladan al núcleo celular e interactúan con regiones del ADN conocidas como elementos de respuesta hormonal (HRE) o se unen a un sitio de unión de otro factor de transcripción. Se ha demostrado que el receptor de estrógeno (RE) transactiva otros factores de transcripción de esta manera, a pesar de la ausencia de un elemento de respuesta al estrógeno (ERE) en la región promotora proximal del gen. Los RE y los receptores de progesterona (RP) son generalmente activadores de genes, con un aumento del ARNm y la subsiguiente síntesis de proteínas tras la exposición hormonal.
Los cerebros masculinos y femeninos difieren en la distribución de los receptores de estrógenos, y esta diferencia es una consecuencia irreversible de la exposición neonatal a los esteroides. Los receptores de estrógenos (y los de progesterona) se encuentran principalmente en las neuronas del hipotálamo anterior y mediobasal, en particular:
- el área preóptica (donde se encuentran las neuronas de la LHRH)
- el núcleo periventricular (donde se encuentran las neuronas de la somatostatina)
- el hipotálamo ventromedial (que es importante para el comportamiento sexual).
Esteroides gonadales en la vida neonatal de las ratas
En la vida neonatal, los esteroides gonadales influyen en el desarrollo del hipotálamo neuroendocrino. Por ejemplo, determinan la capacidad de las hembras para mostrar un ciclo reproductivo normal, y de los machos y hembras para mostrar comportamientos reproductivos adecuados en la vida adulta.
- Si a una rata hembra se le inyecta una vez testosterona en los primeros días de vida postnatal (durante el «periodo crítico» de influencia de los esteroides sexuales), el hipotálamo se masculiniza de forma irreversible; la rata adulta será incapaz de generar una oleada de LH en respuesta al estrógeno (una característica de las hembras), pero será capaz de mostrar comportamientos sexuales masculinos (montar a una hembra sexualmente receptiva).
- Por el contrario, una rata macho castrada justo después de nacer se feminizará, y el adulto mostrará comportamientos sexuales femeninos en respuesta a los estrógenos (receptividad sexual, lordosis}.
Los andrógenos en los primates
En los primates, la influencia del desarrollo de los andrógenos es menos clara, y las consecuencias son menos completas. El «marimacho» en las niñas podría reflejar los efectos de los andrógenos en el cerebro fetal, pero muchos creen que el sexo de la crianza durante los primeros 2-3 años es el determinante más importante de la identidad de género, porque durante esta fase el estrógeno o la testosterona tendrán efectos permanentes en el cerebro femenino o masculino, influyendo tanto en la heterosexualidad como en la homosexualidad.
La paradoja es que los efectos masculinizantes de la testosterona están mediados por el estrógeno. Dentro del cerebro, la testosterona se aromatiza a (estradiol), que es la principal hormona activa para influir en el desarrollo. El testículo humano segrega altos niveles de testosterona desde aproximadamente la octava semana de vida fetal hasta 5-6 meses después del nacimiento (en muchas especies se observa un aumento perinatal similar de la testosterona), un proceso que parece ser la base del fenotipo masculino. El estrógeno de la circulación materna es relativamente ineficaz, en parte debido a los altos niveles circulantes de proteínas de unión a esteroides en el embarazo.
Otras influencias sobre el desarrollo hipotalámico
Los esteroides sexuales no son las únicas influencias importantes sobre el desarrollo hipotalámico; en particular, el estrés prepuberal en los primeros años de vida determina la capacidad del hipotálamo adulto para responder a un factor de estrés agudo. A diferencia de los receptores de esteroides gonadales, los receptores de glucocorticoides están muy extendidos por todo el cerebro; en el núcleo paraventricular, median en el control de retroalimentación negativa de la síntesis y secreción de CRF, pero en otros lugares su papel no se conoce bien.
Efectos del envejecimiento en el hipotálamo
Estudios en ratones hembra han demostrado que tanto el núcleo supraóptico (SON) como el núcleo paraventricular (PVN) pierden aproximadamente un tercio de las células inmunorreactivas de IGF-1R con el envejecimiento normal. Además, los ratones viejos con restricción calórica (CR) perdieron un mayor número de células no inmunorreactivas de IGF-1R mientras mantenían recuentos similares de células inmunorreactivas de IGF-1R en comparación con los ratones viejos-Al. En consecuencia, los ratones Old-CR muestran un mayor porcentaje de células inmunorreactivas de IGF-1R que reflejan una mayor sensibilidad hipotalámica al IGF-1 en comparación con los ratones que envejecen normalmente.
Véase también
- Eje HPA
- Sistema hipofisario hipotálamo
- Sistema adrenal hipofisario hipotálamo
- Lesiones del hipotálamo
- Hermano anterior del cerebro
- Neuroendocrinología
- Área preóptica
Imágenes adicionales
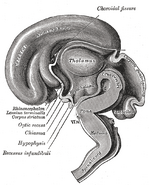
Añadir una foto a esta galería
- http://www.cancer.gov/Templates/db_alpha.aspx?CdrID=46359
- http://www.sci.uidaho.edu/med532/hypothal.htm
- Diagrama de Núcleos (psicopedagogía.org)
- Diagrama de Núcleos (universe-review.ca)
- Diagrama de Núcleos (utdallas.edu)
- Salvo que se especifique otra cosa en la tabla, la referencia es: Guyton Eight Edition
- Walter F., PhD. Boro. Medical Physiology: A Cellular And Molecular Approaoch, Elsevier/Saunders. Página 840
- http://thalamus.wustl.edu/course/hypoANS.html
- http://www.vivo.colostate.edu/hbooks/pathphys/endocrine/hypopit/overview.html
- Theologides A (1976). Metabolitos intermediarios productores de anorexia. Am J Clin Nutr 29 (5): 552-8.
- John Money, ‘The concept of gender identity disorder in childhood and adolescence after 39 years’, Journal of Sex and Marital Therapy 20 (1994): 163-77.
- Romeo, Russell D, Rudy Bellani, Ilia N. Karatsoreos, Nara Chhua, Mary Vernov, Cheryl D. Conrad y Bruce S. McEwen (2005). Stress History and Pubertal Development Interact to Shape Hypothalamic-Pituitary-Adrenal Axis Plasticity. Endocrinología 147 (4): 1664-1674.
- Saeed O,Yaghmaie F,Garan SA,Gouw AM,Voelker MA,Sternberg H, Timiras PS. (2007). Las células inmunorreactivas del receptor del factor de crecimiento similar a la insulina 1 se mantienen selectivamente en el hipotálamo paraventricular de ratones con restricción calórica. Int J Dev Neurosci 25 (1): 23-8.
- Yaghmaie F, Saeed O, Garan SA, Voelker MA, Gouw AM, Freitag W, Sternberg H, Timiras PS (2006). La pérdida dependiente de la edad de células inmunorreactivas del receptor del factor de crecimiento similar a la insulina en el hipotálamo supraóptico se reduce en los ratones con restricción calórica. Int J Dev Neurosci 24 (7): 431-6.
- F. Yaghmaie, O. Saeed, S.A. Garan, A.M. Gouw, P. Jafar, J. Kaur, S. Nijjar, P.S. Timiras, H. Sternberg, M.A. Voelker (2007). Seguimiento de los cambios en la sensibilidad del IGF-1 hipotalámico con el envejecimiento y la restricción calórica. Gerontología experimental 42 (1-2): 148-149.
Referencias & Bibliografía
Papeles
- Arees, E.A. y Mayer, B, (1967) Anatomical connections between medial and lateral regions of the hypothalamus concerned with food intake, Science 157: 1574-5
- Grossman, S.P. (1960) Eating or drinking elicited by direct adrenergic or cholinergic stimulation of the hypothalamus, Science 132: 301-2.
- Hetherington, A.W. and Ranson, S.W. (1942) The relation of various hypothalamic lesions to adiposity in the rat, 76: 475-99.
Material adicional
Papeles
- Google Scholar
- Mapas cerebrales en UCDavis Hipotálamo
- Sistema endocrino e hipotálamo
- AltaResolución Cytoarchitectural Primate Brain Atlases
- The Hypothalamus and Pituitary at endotexts.org
- Diagrama de núcleos (psycheducation.org)
- Diagrama de núcleos (universe-review.ca)
- Diagrama de núcleos (utdallas.edu)
|
v-d-e
Cerebro humano: diencéfalo (TA A14.1.08, GA 9.807) |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Epitálamo |
|
Superficie
|
||||||||||||||
| Tálamo |
|
Superficie
|
Materia gris/
núcleos |
|||||||||||||
| Hipotálamo |
|
|||||||||||||||
| Subtálamo |
Núcleo subtalámico – Zona incerta |
|||||||||||||||
| Sistema ventricular: Tercer ventrículo |
recesos: (receso óptico, receso infundibular, receso suprapineal, receso pineal) Surco hipotalámico – Tela corioidea del tercer ventrículo Órgano subfornical Aperturas: Interventricular/Monro Comisura posterior |
|||||||||||||||
|
v-d-e
Anatomía humana, sistema endocrino: glándulas endocrinas |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ejes hipotalámico/ hipófisis |
|
|
|
|
||||||||||||
| Glándula pineal |
Pinealocitos – Cuerpos arenosos |
|||||||||||||||
| Islotes de páncreas |
Célula alfa – célula beta – célula delta – célula PP – célula épsilon |
|||||||||||||||
Cerebro humano: Sistema límbico
Amígdala – Giro cingulado – Giro fornicario – Hipocampo – Hipotálamo – Cuerpo mamario – Núcleo accumbens – Corteza orbitofrontal – Giro parahipocampal