Posuzování | Biopsychologie | Srovnávací | Kognitivní | Vývojová | Jazyk | Individuální rozdíly | Osobnost | Filozofie | Sociální |
Metody | Statistika | Klinická | Vzdělávací | Průmyslová | Odborné předměty | Světová psychologie |
Biologická:Behaviorální genetika – Evoluční psychologie – Neuroanatomie – Neurochemie – Neuroendokrinologie -Neurologie – Psychoneuroimunologie – Fyziologická psychologie – Psychofarmakologie(Rejstřík, Osnova)
| Mozog: Hypotalamus | ||
|---|---|---|
 |
||
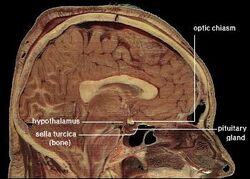
| Uložení lidského hypotalamu | ||
 |
||
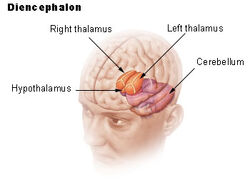
| Dienchephalon | ||
| Latinský | hypothalamus | |
| Grayův | předmět č. 189 812 | |
| Část. z | ||
| Složky | ||
| Arterie | ||
| Vein | ||
| Mozkové informace/UW | hier-358 | |
| MeSH | A08.186.211.730.385.357 | |
Hipotalamus spojuje nervový systém s endokrinním systémem prostřednictvím hypofýzy (podvěsku mozkového). Hypotalamus (z řeckého ὑποθαλαμος = pod thalamem) se nachází pod thalamem, těsně nad mozkovým kmenem. Tato žláza zabírá větší část ventrální oblasti diencephala. Nachází se v mozku všech savců. U člověka má velikost zhruba mandle.
Hypotalamus reguluje některé metabolické procesy a další činnosti autonomního nervového systému. Syntetizuje a vylučuje neurohormony, často nazývané hypotalamické uvolňující hormony, a ty zase stimulují nebo inhibují sekreci hormonů hypofýzy.
Hypotalamus řídí tělesnou teplotu, hlad, žízeň, únavu, hněv a cirkadiánní cykly.
- Vstupy
- Čichové podněty
- Podněty přenášené krví
- Steroidy
- Nervové vstupy
- Jádra
- Výstupy
- Nervové projekce
- Endokrinní hormony
- Řízení příjmu potravy
- Pohlavní dimorfismus
- Reakce na ovariální steroidy
- Gonadální steroidy v neonatálním životě potkanů
- Androgeny u primátů
- Další vlivy na vývoj hypotalamu
- Vliv stárnutí na hypotalamus
- Viz také
- Další obrázky
- Literatura & Bibliografie
- Dokumenty
- Další materiály
- Dokumenty
Vstupy
Hypotalamus je velmi složitá oblast v mozku člověka a i malá jádra v hypotalamu se podílejí na mnoha různých funkcích. Například paraventrikulární jádro obsahuje neurony oxytocinu a vazopresinu (nazývaného také antidiuretický hormon), které se promítají do zadní hypofýzy, ale obsahuje také neurony regulující sekreci ACTH a TSH (které se promítají do přední hypofýzy), žaludeční reflexy, mateřské chování, krevní tlak, krmení, imunitní reakce a teplotu.
Hypotalamus koordinuje mnoho hormonálních a behaviorálních cirkadiánních rytmů, složité vzorce neuroendokrinních výstupů, komplexní homeostatické mechanismy a mnoho důležitých chování. Hypotalamus proto musí reagovat na mnoho různých signálů, z nichž některé jsou generovány externě a některé interně. Je tedy bohatě propojen s mnoha částmi CNS, včetně retikulární formace mozkového kmene a autonomních zón, limbického předního mozku (zejména amygdaly, septa, diagonálního pásu Broca a čichových bulbů a mozkové kůry).
Hypotalamus reaguje na:
- Světlo: délku dne a fotoperiodu pro regulaci cirkadiánních a sezónních rytmů
- Olfaktorické podněty, včetně feromonů
- Steroidy, včetně gonadálních steroidů a kortikosteroidů
- Neurálně přenášené informace pocházející zejména ze srdce, žaludku, a reprodukčního traktu
- Autonomní vstupy
- Krevní podněty, včetně leptinu, ghrelinu, angiotenzinu, inzulínu, hormonů hypofýzy, cytokinů, plazmatických koncentrací glukózy a osmolarity atd
- Stres
- Invaze mikroorganismů zvýšením tělesné teploty, resetování termostatu těla směrem nahoru.
Čichové podněty
Čichové podněty jsou u mnoha druhů důležité pro rozmnožování a neuroendokrinní funkce. Například pokud je březí myš vystavena moči „cizího“ samce v kritickém období po souloži, pak těhotenství selže (Bruceův efekt). Samice myši si tak během soulože vytvoří přesnou „čichovou paměť“ na svého partnera, která přetrvává několik dní. feromonální signály napomáhají synchronizaci říje u mnoha druhů; u žen může synchronizovaná menstruace také vznikat na základě feromonálních signálů, ačkoli o úloze feromonů u lidí někteří pochybují.
Podněty přenášené krví
Peptidové hormony mají významný vliv na hypotalamus, a aby tak mohly učinit, musí se vyhnout hematoencefalické bariéře. Hypotalamus je částečně ohraničen specializovanými oblastmi mozku, které postrádají účinnou hematoencefalickou bariéru; endotel kapilár v těchto místech je fenestrovaný, aby umožnil volný průchod i velkých proteinů a dalších molekul. Některá z těchto míst jsou místy neurosekrece – neurohypofýza a střední eminence. Jiná místa jsou však místy, kde mozek odebírá vzorky složení krve. Dvě z těchto míst, subfornikální orgán a OVLT (organum vasculosum of the lamina terminalis), jsou tzv. cirkumventrikulární orgány, kde jsou neurony v těsném kontaktu s krví i mozkomíšním mokem. Tyto struktury jsou hustě vaskularizované a obsahují osmoreceptivní a sodík-receptivní neurony, které řídí pití, uvolňování vazopresinu, vylučování sodíku a chuť k příjmu sodíku. Obsahují také neurony s receptory pro angiotenzin, atriální natriuretický faktor, endotelin a relaxin, z nichž každý je důležitý pro regulaci rovnováhy tekutin a elektrolytů. Neurony v OVLT a SFO se promítají do supraoptického jádra a paraventrikulárního jádra a také do preoptických hypotalamických oblastí. Obvodové orgány mohou být také místem působení interleukinů, které vyvolávají horečku i sekreci ACTH prostřednictvím účinků na paraventrikulární neurony.
Není jasné, jak všechny peptidy, které ovlivňují hypotalamickou aktivitu, získávají potřebný přístup. V případě prolaktinu a leptinu existují důkazy o aktivním vychytávání v choroidálním plexu z krve do mozkomíšního moku. Některé hypofyzární hormony mají negativní zpětnou vazbu na hypotalamickou sekreci; například růstový hormon se vrací zpět do hypotalamu, ale není jasné, jak se dostává do mozku. Existují také důkazy o centrálním působení prolaktinu a TSH.
Steroidy
Hypotalamus obsahuje neurony citlivé na gonadální steroidy a glukokortikoidy – (steroidní hormony nadledvin, uvolňované v reakci na ACTH). Obsahuje také specializované neurony citlivé na glukózu (v arcuate nucleus a ventromediálním hypotalamu), které jsou důležité pro chuť k jídlu. Preoptická oblast obsahuje termosenzitivní neurony; ty jsou důležité pro sekreci TRH.
Nervové vstupy
Hypotalamus dostává mnoho vstupů z mozkového kmene; zejména z jádra solitárního traktu, locus coeruleus a ventrolaterální dřeně. Některými z těchto drah je zprostředkována sekrece oxytocinu v reakci na sání nebo vaginocervikální stimulaci; jinými je zprostředkována sekrece vazopresinu v reakci na kardiovaskulární podněty vycházející z chemoreceptorů v karotickém sinu a aortálním oblouku a z receptorů nízkotlakého objemu síní. U potkana způsobuje stimulace pochvy také sekreci prolaktinu, což vede k pseudobřezosti po neplodném páření. U králíka vyvolává koitus reflexní ovulaci. U ovcí může stimulace děložního hrdla v přítomnosti vysokých hladin estrogenů vyvolat mateřské chování u panenské ovce. Všechny tyto účinky jsou zprostředkovány hypotalamem a informace jsou přenášeny především míšními drahami, které se přenášejí v mozkovém kmeni. Stimulace bradavek stimuluje uvolňování oxytocinu a prolaktinu a potlačuje uvolňování LH a FSH.
Kardiovaskulární podněty jsou přenášeny bloudivým nervem, ale bloudivý nerv přenáší také různé viscerální informace, včetně například signálů vznikajících při roztažení žaludku k potlačení krmení. Tyto informace se opět dostávají do hypotalamu prostřednictvím relé v mozkovém kmeni.
Jádra
Mezi hypotalamická jádra patří následující:
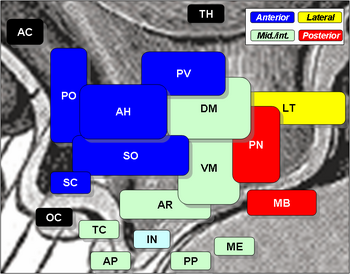
Hypotalamická jádra
| Region | Area | Nukleus | Funkce |
| Přední | Mediální | Mediální preoptické jádro |
|
| supraoptické jádro (SO) |
|
||
| paraventrikulární jádro (PV) |
|
||
| přední hypotalamické jádro (AH) |
|
||
| Suprachiasmatické jádro (SC) |
|
||
| Laterální | Laterální preoptické jádro | ||
| Laterální jádro (LT) |
|
||
| Část supraoptického jádra (SO) |
|
||
| Tuberální | Mediální | Dorzomediální hypotalamické jádro (DM) |
|
| Ventromediální jádro (VM) |
. |
||
Arkuátní jádro (AR)
|
|||
| Laterální | . Laterální jádro (LT) |
|
|
| Laterální tuberální jádro | |||
| Posteriorní | Mediální | Mamilární jádra (součást mamilárních tělísek) (MB) |
|
| zadní jádro (PN) |
|
||
| Laterální | Laterální jádro (LT) |
Viz též: ventrolaterální preoptické jádro
Výstupy
Výstupy hypotalamu lze rozdělit do dvou kategorií: nervové projekce a endokrinní hormony.
Nervové projekce
Většina systémů vláken hypotalamu probíhá obousměrně (obousměrně).
- Projekce do oblastí kaudálně od hypotalamu jdou přes mediální svazek předního mozku, mammillotegmentální trakt a dorzální podélný fascikulus.
- Projekce do oblastí rostrálně od hypotalamu jsou vedeny mammillothalamickým traktem, fornixem a terminálním striem.
Endokrinní hormony
Hypotalamus ovlivňuje endokrinní systém a řídí emoční chování, například hněv a sexuální aktivitu. Většina hypotalamem vytvořených hormonů je distribuována do hypofýzy prostřednictvím hypofyzárního portálního systému. Hypotalamus udržuje homeostázu, což zahrnuje regulaci krevního tlaku, srdeční frekvence a teploty.
Primární hypotalamické hormony jsou:
| Název | Jiné názvy | Zkratky | Umístění | Funkce |
|---|---|---|---|---|
| Corticotropin-releasing hormone | Corticotropin-releasing factor, Corticoliberin | CRH, CRF | parvocelulární neuroendokrinní neurony v paraventrikulárním jádru | s vazopresinem, stimuluje přední hypofýzu k sekreci ACTH |
| Dopamin | Prolaktin-inhibiční hormon | DA, PIH | neuroendokrinní neurony arcuate nucleus | inhibuje sekreci prolaktinu z přední hypofýzy |
| Hormon uvolňující gonadotropin | Hormon uvolňující luteinizační hormon | GnRH, LHRH | neuroendokrinní neurony v mediálních preoptických a arkuátních jádrech | stimuluje přední hypofýzu k sekreci LH a FSH |
| hormon uvolňující růstový hormon | faktor uvolňující růstový hormon, somatokrinin | GHRH, GHRF, GRF | neuroendokrinní neurony arcuate nucleus | stimuluje přední hypofýzu k sekreci růstového hormonu |
| Melatonin | suprachiasmatické jádro | |||
| Somatostatin | Růstový hormon-inhibiční hormon, Somatotropin release-inhibiting factor | SS, GHIH, SRIF | neuroendokrinní neurony periventrikulárního jádra | inhibuje sekreci růstového hormonu z přední hypofýzy |
| Thyrotropin-releasing hormone | Thyrotropin-releasing factor, Thyroliberin, Protirelin | TRH, TRF | parvocelulární neuroendokrinní neurony v paraventrikulárních a předních hypotalamických jádrech | stimuluje přední hypofýzu k sekreci TSH |
Viz též: Hypokretin
Řízení příjmu potravy
Krajní laterální část ventromediálního jádra hypotalamu je zodpovědná za řízení příjmu potravy. Stimulace této oblasti způsobuje zvýšený příjem potravy. Bilaterální léze této oblasti způsobuje úplné zastavení příjmu potravy. Mediální část jádra má řídící účinek na laterální část. Bilaterální léze mediální části ventromediálního jádra způsobuje hyperfagii a obezitu zvířete. Další léze laterální části ventromediálního jádra u téhož zvířete způsobí úplné zastavení příjmu potravy.
Existují různé hypotézy týkající se této regulace:
- Lipostatická hypotéza – tato hypotéza tvrdí, že tuková tkáň produkuje humorální signál, který je úměrný množství tuku a působí na hypotalamus tak, že snižuje příjem potravy a zvyšuje výdej energie. Je zřejmé, že na hypotalamus působí hormon leptin, který snižuje příjem potravy a zvyšuje výdej energie.
- Gutpeptidová hypotéza – gastrointestinální hormony jako Grp, glukagony, CCK a další údajně inhibují příjem potravy. Potrava vstupující do gastrointestinálního traktu vyvolává uvolňování těchto hormonů, které působí na mozek a vyvolávají pocit sytosti. Mozek obsahuje receptory CCK-A i CCK-B.
- Glukostatická hypotéza – aktivita centra sytosti ve ventromediálních jádrech se pravděpodobně řídí využitím glukózy v neuronech. Bylo postulováno, že když je jejich utilizace glukózy nízká, a tudíž když je rozdíl arteriovenózní glukózy v krvi přes ně nízký, aktivita přes neurony se snižuje. Za těchto podmínek je aktivita centra výživy nekontrolovaná a jedinec pociťuje hlad. Příjem potravy se rychle zvyšuje intraventrikulárním podáním 2-deoxyglukózy, a proto se snižuje utilizace glukózy v buňkách.
- Thermostatická hypotéza – podle této hypotézy snížení tělesné teploty pod danou stanovenou hodnotu stimuluje chuť k jídlu, zatímco zvýšení nad stanovenou hodnotu chuť k jídlu potlačuje.
Pohlavní dimorfismus
Několik hypotalamických jader je pohlavně dimorfních, tj. existují jasné rozdíly ve struktuře i funkci mezi samci a samicemi.
Některé rozdíly jsou patrné i v hrubé neuroanatomii: nejpozoruhodnější je pohlavně dimorfní jádro v preoptické oblasti, které je přítomno pouze u samců. Většinu rozdílů však představují jemné změny v konektivitě a chemické citlivosti jednotlivých souborů neuronů.
Důležitost těchto změn lze rozpoznat podle funkčních rozdílů mezi samci a samicemi. Například vzorec sekrece růstového hormonu je pohlavně dimorfní, a to je jeden z důvodů, proč jsou u mnoha druhů dospělí samci mnohem větší než samice.
Reakce na ovariální steroidy
Další nápadné funkční dimorfismy jsou v behaviorálních reakcích na ovariální steroidy dospělých jedinců. Muži a ženy reagují na ovariální steroidy odlišně, částečně proto, že exprese neuronů citlivých na estrogeny v hypotalamu je pohlavně dimorfní, tj. estrogenové receptory jsou exprimovány v různých sadách neuronů.
Estrogen a progesteron mohou ovlivňovat genovou expresi v určitých neuronech nebo vyvolávat změny v membránovém potenciálu buněk a aktivaci kináz, což vede k různým negenomovým buněčným funkcím. Estrogen a progesteron se vážou na jim příbuzné receptory jaderných hormonů, které se přemisťují do buněčného jádra a interagují s oblastmi DNA známými jako elementy hormonální odpovědi (HRE) nebo se vážou na vazebné místo jiného transkripčního faktoru. Bylo prokázáno, že estrogenový receptor (ER) tímto způsobem transaktivuje jiné transkripční faktory, přestože v proximální promotorové oblasti genu chybí element estrogenové odpovědi (ERE). ER a progesteronové receptory (PR) jsou obecně genovými aktivátory se zvýšenou mRNA a následnou syntézou proteinů po expozici hormonům.
Mozky mužů a žen se liší v distribuci estrogenových receptorů a tento rozdíl je nevratným důsledkem neonatální expozice steroidům. Estrogenové receptory (a receptory progesteronu) se nacházejí především v neuronech v předním a mediobazálním hypotalamu, zejména:
- v preoptické oblasti (kde se nacházejí neurony LHRH)
- v periventrikulárním jádru (kde se nacházejí neurony somatostatinu)
- ve ventromediálním hypotalamu (který je důležitý pro sexuální chování).
Gonadální steroidy v neonatálním životě potkanů
V neonatálním životě ovlivňují gonadální steroidy vývoj neuroendokrinního hypotalamu. Určují například schopnost samic vykazovat normální reprodukční cyklus a samců a samic vykazovat vhodné reprodukční chování v dospělosti.
- Pokud je samici potkana v prvních dnech postnatálního života (během „kritického období“ vlivu pohlavních steroidů) jednorázově aplikován testosteron, hypotalamus se nevratně maskulinizuje; dospělý potkan nebude schopen generovat vzestup LH v reakci na estrogen (což je charakteristické pro samice), ale bude schopen projevovat samčí sexuální chování (osedlat sexuálně vnímavou samici).
- Naproti tomu samec potkana vykastrovaný těsně po narození bude feminizován a dospělý jedinec bude vykazovat samičí sexuální chování v reakci na estrogen (sexuální receptivita, lordóza}.
Androgeny u primátů
U primátů je vývojový vliv androgenů méně jasný a důsledky jsou méně úplné. ‚Tomboyismus‘ u dívek by mohl odrážet účinky androgenů na mozek plodu, ale pohlaví při výchově během prvních 2-3 let je podle mnohých nejdůležitějším faktorem určujícím pohlavní identitu, protože během této fáze buď estrogen, nebo testosteron trvale ovlivní buď ženský, nebo mužský mozek, což má vliv jak na heterosexualitu, tak na homosexualitu.
Paradoxem je, že maskulinizující účinky testosteronu jsou zprostředkovány estrogenem. V mozku je testosteron aromatizován na (estradiol), který je hlavním aktivním hormonem pro vývojové vlivy. Lidské varle vylučuje vysoké hladiny testosteronu přibližně od 8. týdne života plodu do 5-6 měsíců po narození (podobný perinatální nárůst testosteronu je pozorován u mnoha živočišných druhů), což je proces, který je zřejmě základem mužského fenotypu. Estrogen z mateřské cirkulace je relativně neúčinný, částečně kvůli vysokým cirkulujícím hladinám proteinů vážících steroidy v těhotenství.
Další vlivy na vývoj hypotalamu
Pohlavní steroidy nejsou jediným důležitým vlivem na vývoj hypotalamu; zejména pre-pubertální stres v raném věku určuje schopnost dospělého hypotalamu reagovat na akutní stresor. Na rozdíl od receptorů pro gonadální steroidy jsou glukokortikoidní receptory velmi rozšířené v celém mozku; v paraventrikulárním jádře zprostředkovávají negativní zpětnovazební kontrolu syntézy a sekrece CRF, ale jinde není jejich role dobře známa.
Vliv stárnutí na hypotalamus
Studie na myších samicích ukázaly, že supraoptické jádro (SON) i paraventrikulární jádro (PVN) ztrácejí při normálním stárnutí přibližně třetinu imunoreaktivních buněk IGF-1R. Také staré myši s omezeným příjmem kalorií (CR) ztratily větší počet neimunoreaktivních buněk IGF-1R při zachování podobného počtu imunoreaktivních buněk IGF-1R ve srovnání se starými myšmi. V důsledku toho vykazují myši Old-CR vyšší procento imunoreaktivních buněk IGF-1R, což odráží zvýšenou citlivost hypotalamu na IGF-1 ve srovnání s normálně stárnoucími myšmi.
Viz také
- HPA osa
- Hypothalamo hypofyzární systém
- Hypothalamo hypofyzární adrenální systém
- Hypothalamus léze
- Mediální přední mozkový svazek
- Neuroendokrinologie
- Preoptická oblast
.
Další obrázky
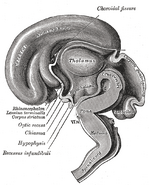
Přidat fotografii do této galerie
- http://www.cancer.gov/Templates/db_alpha.aspx?CdrID=46359
- http://www.sci.uidaho.edu/med532/hypothal.htm
- Schéma jádra (psycheducation.org)
- Diagram jader (universe-review.ca)
- Diagram jader (utdallas.edu)
- Pokud není v tabulce uvedeno jinak, pak je ref: Guyton Eight Edition
- Walter F., PhD. Bór. Lékařská fyziologie: A Cellular And Molecular Approaoch, Elsevier/Saunders. Strana 840
- http://thalamus.wustl.edu/course/hypoANS.html
- http://www.vivo.colostate.edu/hbooks/pathphys/endocrine/hypopit/overview.html
- Theologides A (1976). Anorexii produkující intermediární metabolity. Am J Clin Nutr 29 (5): 552-8.
- John Money, „The concept of gender identity disorder in childhood and adolescence after 39 years“ (Koncept poruchy pohlavní identity v dětství a dospívání po 39 letech), Journal of Sex and Marital Therapy 20 (1994): 163-77.
- Romeo, Russell D, Rudy Bellani, Ilia N. Karatsoreos, Nara Chhua, Mary Vernov, Cheryl D. Conrad a Bruce S. McEwen (2005). Stresová historie a pubertální vývoj vzájemně ovlivňují plasticitu osy hypotalamus-hypofýza-nadledviny. Endocrinology 147 (4): 1664-1674.
- Saeed O,Yaghmaie F,Garan SA,Gouw AM,Voelker MA,Sternberg H, Timiras PS. (2007). Imunoreaktivní buňky receptoru pro inzulínu podobný růstový faktor-1 jsou selektivně udržovány v paraventrikulárním hypotalamu myší s omezeným kalorickým příjmem. Int J Dev Neurosci 25 (1): 23-8.
- Yaghmaie F, Saeed O, Garan SA, Voelker MA, Gouw AM, Freitag W, Sternberg H, Timiras PS (2006). Úbytek imunoreaktivních buněk receptoru pro růstový faktor podobný inzulínu-1 v supraoptickém hypotalamu závislý na věku je snížen u myší s omezeným přísunem kalorií. Int J Dev Neurosci 24 (7): 431-6.
- F. Yaghmaie, O. Saeed, S.A. Garan, A.M. Gouw, P. Jafar, J. Kaur, S. Nijjar, P.S. Timiras, H. Sternberg, M.A. Voelker (2007). Sledování změn citlivosti hypotalamu na IGF-1 se stárnutím a kalorickou restrikcí. Experimental Gerontology 42 (1-2): 148-149.
Literatura & Bibliografie
Dokumenty
- Arees, E.A. a Mayer, B, (1967) Anatomical connections between medial and lateral regions of the hypothalamus concerned with food intake (Anatomická spojení mezi mediálními a laterálními oblastmi hypotalamu, které se týkají příjmu potravy), Science 157: 1574-5
- Grossman, S.P. (1960) Elicited or drinking by direct adrenergic or cholinergic stimulation of the hypothalamus, Science 132: 301-2.
- Hetherington, A.W. and Ranson, S.W. (1942) The relation of various hypothalamic lesions to adiposity in the rat, 76: 475-99. (Vztah různých hypotalamických lézí k tučnosti u potkana).
Další materiály
Dokumenty
- Google Scholar
.
- BrainMaps at UCDavis Hypothalamus
- Endokrinní systém a hypothalamus
- Vysoká-Resolution Cytoarchitectural Primate Brain Atlases
- Hypotalamus a hypofýza na endotextech.org
- Diagram jader (psycheducation.org)
- Diagram jader (universe-review.ca)
- Diagram jader (utdallas.edu)
|
v-d-e
Lidský mozek: diencephalon (TA A14.1.08, GA 9.807) |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Epithalamus |
|
Povrch
|
||||||||||||||
| Thalamus |
|
Povrch
|
Šedá hmota/
jádra |
|||||||||||||
| Hypothalamus |
|
|||||||||||||||
| Subthalamus |
Subthalamické jádro – Zona incerta |
|||||||||||||||
| Komorový systém: Třetí komora |
recesy: (Optický reces, Infundibulární reces, Suprapineální reces, Pineální reces) Hypothalamický sulcus – Tela chorioidea třetí komory Subfornální orgán Apertury: Interventrikulární/Monro Zadní komisura |
|||||||||||||||
|
v-d-e
Anatomie člověka, endokrinní systém: endokrinní žlázy |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hypotalamická/ hypofyzární osa |
|
|
|
|
||||||||||||
| Pineální žláza |
Pinealocyty – Corpora arenacea |
|||||||||||||||
| Islets of pankreatu |
Alfa buňka – Beta buňka – Delta buňka – PP buňka – Epsilon buňka |
|||||||||||||||
Lidský mozek: Limbický systém
Amygdala – Cingulární gyrus – Smilný gyrus – Hipokampus – Hypotalamus – Mamilární tělísko – Nucleus accumbens – Orbitofrontální kůra – Parahipokampální gyrus
.