Úvod
Šlechtitelé i výzkumníci se zajímají o hybridizaci a introgresi mezi odlišnými genotypy jako o potenciální faktor vedoucí k ekologické divergenci potomstva (Rieseberg et al., 2007; Arnold et al., 2012). Naopak hybridizace může někdy zahrnovat sníženou hybridní fitness, jako je slabost a/nebo sterilita v F1 a pozdějších generacích, a to i v případech hybridizace mezi příslušníky stejného druhu.
Snížená hybridní životaschopnost a/nebo plodnost segregující v F2 nebo pozdějších generacích se označuje jako hybridní porucha (HB), s níž jsou nutně spojeny recesivní alely. Tato reprodukční bariéra je u rostlin i živočichů pozorována již dlouhou dobu (Dobzhansky, 1970; Grant, 1971). Mnoho genetických analýz této bariéry odhalilo, že se uskutečňuje komplementárním efektem mezi lokusy a/nebo mezi lokusy s diferencovanými alelami, běžně nazývaným Bateson-Dobzhansky-Mullerova (BDM) inkompatibilita (Rieseberg a Willis, 2007). HB nutně zahrnuje vnitřní postzygotické reprodukční bariéry, jako je hybridní inviabilita (včetně slabosti, nekrózy a chlorózy) a hybridní sterilita (u samců, samic nebo obou gamet); někteří výzkumníci proto nemusí rozlišovat HB od inviability a sterility u potomstva F1. Nicméně se domnívám, že HB je vhodnou klasifikací reprodukčních bariér, protože implikuje jejich základní genetický základ (tj. zapojení recesivních alel).
Nedávné studie využívající modelový druh rostliny Arabidopsis umožnily lepší pochopení HB, pokud jde o její molekulární mechanismy (Vaid a Laitinen, 2019). Naopak, ačkoli je rýže (druh Oryza) modelovou plodinou, současné znalosti genetického základu HB zůstávají u tohoto druhu omezené, pravděpodobně proto, že HB nepředstavuje reprodukční překážku u F1 hybridů, u nichž se očekává vyšší produkce zrna ve srovnání s rodičovskými inbredními liniemi, a pravděpodobně proto, že méněcenné rostliny, které segregují v F2 a pozdějších generacích, mohou být snadno selektovány na základě fenotypu ze šlechtitelské populace. Za tímto omezením mohou být také obtíže při genetickém mapování způsobené recesivní dědičností.
Přibližuji zde HB v kříženích rýže, přičemž se odvolávám na informace poskytnuté studiemi Arabidopsis, a diskutuji o tom, jak se s HB zachází při šlechtění rýže.
Genetický základ hybridního rozpadu u rýže
Předpokládá se, že kultivovaná rýže, O. sativa japonica a indica, tvoří druhový komplex se svým domnělým prapředkem (O. rufipogon) a divokými druhy; při křížení mezi nimi jsou však pozorovány četné reprodukční bariéry, včetně HB (Oka, 1988; Vaughan et al.),
Pokud je mi známo, Dr. Oka jako první popsal genetický základ HB u mezidruhových křížení japoniky a indiky, který byl vysvětlen dvěma komplementárními geny (Oka, 1957). Odpovědné geny však nebyly zmapovány na chromozomech, protože v té době nebyly k dispozici molekulární markery.
U hybridů rýže byla HB často popisována u mezidruhových (O. sativa ssp. japonica × ssp. indica) a mezidruhových (O. sativa × O. nivara a O. sativa × O. glumaepatula) křížení (Oka, 1957; Sato a Morishima, 1988; Wu et al., 1995; Li et al., 1997; Fukuoka et al., 1998, 2005; Kubo a Yoshimura, 2002, 2005; Matsubara et al., 2007a,b, 2015; Yamamoto et al., 2007; Ichitani et al., 2012;pro mezidruhové křížení; Sobrizal et al., 2001; Miura et al., 2008 pro mezidruhové křížení) (tabulka 1). Většina těchto případů HB byla způsobena inkompatibilitou dvou lokusů BDM a recesivními alelami, přičemž 1/16 potomstva F2, které bylo homozygotní pro recesivní alely v obou lokusech, vykazovalo fenotyp HB; avšak 4/16 potomstva, které bylo heterozygotní pouze v jednom lokusu, vykazovalo fenotyp HB v závislosti na kombinaci křížení (tabulka 1 a obrázek 1A). U zpětně zkřížených hybridů byl jeden lokus již fixován alelami od rekurentního rodiče; proto 1/4 potomstva BCnF2 vykazovala HB fenotyp (tabulka 1 a obrázek 1B).

Tabulka 1. Genetický základ hybridního rozpadu zaznamenaného u křížení rýže.
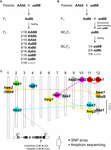
Obrázek 1. Znázornění hybridního rozpadu. Genetický základ hybridního rozpadu (HB). (A) Schematické znázornění genotypu HB, který segreguje v potomstvu F2. Pokud jsou rodičovské genotypy AAbb a aaBB, 1/16 potomstva F2 bez dominantní alely, která je označena šedým znakem (tj. aabb), vykazuje sníženou životaschopnost a/nebo plodnost, ačkoli ostatní genotypy jsou normální. V některých kříženích rýže potomci F2 s pouze jednou dominantní alelou (tj. Aabb, aaBb) rovněž vykazují sníženou životaschopnost a/nebo plodnost. (B) Schematické znázornění genotypu HB, který segreguje v potomstvu BCnF2. Dále 1/4 potomstva BCnF2 vykazuje sníženou životaschopnost a/nebo plodnost. (C) Chromozomální umístění genů, které jsou základem HB u kříženců rýže. Geny jsou přibližně zmapovány na základě výsledků mapování genů v jednotlivých studiích. Soubor komplementárních genů je spojen přerušovanou čarou. Komplementární geny s hbd1 a hwf1 nebyly zaznamenány. Je uveden příklad funkčního nukleotidového polymorfismu způsobujícího HB, který publikovali Yamamoto et al. (2010), a tyto informace nám umožní provést průzkum pomocí SNP array nebo amplikonového sekvenování.
Fenotyp HB u hybridů rýže se liší podle kombinace křížení. Například genotyp hbd2/hbd2 hbd3/hbd3 vykazuje slabost, ale žádnou zjevnou sterilitu semen. Genotyp hbd4/hbd4 hbd5/hbd5 však vykazuje slabost i sterilitu semen (Matsubara et al., 2007a, 2015; Yamamoto et al., 2007; tab. 1). Existují také komplikované případy, kdy se vyskytují HB vykazující slabost i sterilitu semen (genotyp hwe1/hwe1 hwe2/hwe2) nebo HB vykazující pouze sterilitu semen (hsa1/hsa1 hsa2/hsa2 hsa3/hsa3), a to i přes stejnou kombinaci křížení, tj. křížení Asominori × IR24 (Kubo a Yoshimura, 2002, 2005; tab. 1). Vzhledem k tomu, že tyto HB mají nezávislý genetický základ, minimálně 5/64 potomků F2 by v tomto křížení vykazovalo fenotyp HB.
Dosud bylo pomocí DNA markerů v kříženích mezi japonskými a indickými odrůdami rýže (Chrs 7 a 10, Fukuoka a kol.) zmapováno několik sad lokusů odpovědných za HB v určitých genomických oblastech, 1998; Chrs 1 a 12, Wu et al., 1995; Kubo a Yoshimura, 2002; Matsubara et al., 2015; Chrs 6 a 11, Fukuoka et al., 2005; Chrs 8, 9 a 12, Kubo a Yoshimura, 2005; Chrs 2 a 11, Matsubara et al., 2007a; Yamamoto et al., 2007, 2010; Chrs 11 a 12, Ichitani et al., 2012; tabulka 1 a obrázek 1C). Výsledky těchto studií odhalily, že lokusy, které jsou základem HB rýže, jsou v některých kříženích společné, ale v jiných se liší, což naznačuje jejich diverzifikaci v genomech rýže. Například v některých případech alela HB nesená druhem O. nivara, který je blízce příbuzný odrůdám incica, sdílí stejný lokus s odrůdou indica; alela HB nesená druhem O. glumaepatula, který je blízce příbuzný kulturním odrůdám, však nesdílí lokus s žádnou jinou odrůdou (tabulka 1 a obrázek 1C). Je třeba poznamenat, že výše popsané případy HB jsou způsobeny souborem genů s hlavními účinky; existují však i případy způsobené souborem genů s menšími účinky, např. mírně sníženou plodností semen, které nelze při šlechtění rýže přehlížet a které dosud nebyly zjištěny.
Molekulární mechanismy, které jsou základem hybridního rozpadu
Autoimunitní odpověď: Bomblies et al. (2007) jako první uvedli, že autoimunitní odpověď, kterou má gen pro rezistenci k chorobám NB-LRR nebo gen R, je spojena s HB u Arabidopsis při vnitrodruhovém křížení, ačkoli tato autoimunitní odpověď se projevuje hlavně jako nekróza u rostlin F1 a HB u potomstva F2 je zřejmě podmíněna teplotou. U hybridů Arabidopsis bylo popsáno několik linií důkazů HB způsobené autoimunitní reakcí (Alcázar et al., 2009, 2010). Zapojení genů NB-LRR do HB naznačuje, že s HB Arabidopsis může být spojeno více genomických oblastí; tuto myšlenku ostatně podporuje i rozsáhlý průzkum provedený Chae et al. (2014).
Také u hybridů rýže byla popsána HB způsobená autoimunitní reakcí u křížení japonica × indica (Yamamoto et al., 2010). V tomto případě se HB objevuje u rostlin F2, když se gen hybridní poruchy 2 (hbd2), který kóduje kaseinkinázu I a který nese odrůda indica, zkombinuje se shlukem genů R nesených odrůdou japonica. Protože se tyto příčinné alely pravděpodobně chovají jako částečně recesivní, může se jednat o produkty těchto genů (Matsubara et al., 2007a; Yamamoto et al., 2007, 2010).
Reciproké umlčení duplikovaných genů: tento pojem byl poprvé teoreticky navržen jako genetický základ hybridní inkompatibility (Werth a Windham, 1991; Lynch a Force, 2000). V tomto modelu jedna populace ztrácí funkci v jednom lokusu a zachovává ji v druhém, zatímco u druhé populace dochází k opačnému efektu. V důsledku toho 1/16 zygot F2 nemá funkční geny (Lynch a Force, 2000). Dobrým příkladem tohoto jevu je HB u hybridů Arabidopsis, který popsali Bikard a kol (2009) a Vlad a kol (2010). V tomto scénáři byly také popsány případy zahrnující epigenetické umlčení (Agorio et al., 2017; Blevins et al., 2017).
Jelikož u rýže došlo k celogenomové i segmentální duplikaci (Wang et al., 2007; Guo et al., 2019), mohlo by jako příčina hybridní inkompatibility převládat vzájemné umlčení. Dosud však neexistují důkazy o HB způsobené recipročním umlčováním duplikovaných genů u rýže.
Nezávisle na autoimunitní reakci a recipročním umlčování byla identifikována interakce mezi DEAD-box RNA helikázou 18 a jiným genem (MORPHEUS MOLECULE 1 jako pravděpodobný kandidát), která způsobuje HB u hybridů Arabidopsis (Plötner et al., 2017; Vaid et al., 2020). Zajímavé je, že fenotyp HB se mezi generacemi F3 a F4 snižuje, což naznačuje epigenetickou regulaci genové exprese.
U hybridů rýže mezi třemi lokusy hybridní sterility-a (hsa) Kubo et al. nedávno ukázali (2016), že lokus hsa1 se skládá ze dvou genů a že tyto geny kódují protein DUF1618 a nepopsaný protein s určitou podobností s proteinem vázajícím nukleotidy, resp. Molekulární vlastnosti zbývajících komplementárních genů, hsa2 a hsa3, nebyly dosud popsány.
Hybridní zlom v konvenčním šlechtění rýže
Po desetiletí se v konvenčním šlechtění rýže hojně využívala metoda hromadné populace. Při této metodě je po křížení populace F4 nebo F5 vychována samoplozením ve velkém (spíše než metodou jednosemenného sestupu) bez umělé selekce, ale populace rané generace je podrobena přirozené selekci a selekci životaschopnosti (Allard, 1960; podrobněji Ikehashi a Fujimaki, 1980). Při metodě hromadné populace lze očekávat, že mnoho slabých a/nebo sterilních genotypů bude z populace vyřazeno před vytvořením populace pokročilé generace; proto může tato metoda poskytovat omezené informace o HB.
Pro introgresi nebo akumulaci žádoucích znaků od dárce (dárců) do odrůdy šlechtitelé rýže často prováděli zpětné křížení nebo křížení více rodičů, po kterém následovala metoda hromadné populace. Empiricky je již dlouho známo, že tyto metody křížení umožňují zmírnit ztráty kandidátů selekce reprodukčními bariérami, protože tyto metody často snižují segregaci rušivých kombinací alel spojených s HB v hybridním potomstvu. Zmírnění reprodukčních bariér těmito metodami křížení je však nevyhnutelně závislé na HB genotypu rodičů.
Hybridní zlom ve šlechtění založeném na genomice
Publikace referenčních sekvencí genomu plodin a rozvoj technologií sekvenování nové generace urychlily pokrok v molekulárním šlechtění plodin (Kole et al., 2015). V této souvislosti se ve šlechtění plodin uvažuje o genomicky asistovaném šlechtění, jako je genomická selekce založená na genotypech celogenomových markerů DNA (Spindel a Iwata, 2018).
V genomicky asistovaném šlechtění samosprašných plodin se jako referenční populace často používají populace pokročilé generace, jako jsou rekombinantní inbrední linie, z nichž se získávají údaje o genotypu a fenotypu celého genomu. Následně se u potomstva raných generací (např. F2) provádí selekce založená pouze na markerových genotypech. V tomto selekčním schématu je třeba poznamenat, že referenční populace obvykle neposkytuje informace o genomické oblasti pro HB, protože slabá a/nebo sterilní potomstva by měla být v raných generacích po křížení vyřazena. Pokud tedy přijmeme toto selekční schéma, budeme muset opustit některé důležité kandidáty selekce. Případně můžeme vybrat nežádoucí kandidáty tak, že se HB projeví v pozdějších generacích.
Diskuse
Přes omezený počet studií na toto téma se zdá, že lokusy, které jsou základem HB v kříženích rýže, jsou spíše diverzifikované než společné (tabulka 1 a obrázek 1). Jak bylo popsáno výše, informace o rozložení alel spojených s HB mezi rodiči kříženců by měly být nezbytným předpokladem pro šlechtění rýže, zejména pro maximalizaci účinnosti genomicky asistovaného šlechtění. Proto je zapotřebí další detekce HB u kříženců rýže a mapování odpovědných genů v genomu rýže. U hybridů Arabidopsis byly takové údaje o R genech a jejich interagujících genech rozsáhle zkoumány (Alcázar et al., 2010, 2014; Chae et al., 2014). Zpráva o genu hbd2 od Yamamota et al. (2010) u hybridu rýže je dobrým příkladem rozsáhlého průzkumu provedeného pomocí SNP polí nebo amplikonového sekvenování (obr. 1C). Kandidáty na tento typ průzkumu jsou také geny hbd3 a hsa1. Vývoj kritérií, která nám umožní předpovídat HB na základě genomických informací rodičovských linií, bude nakonec sloužit jako důležitý nástroj pro genomicky asistované šlechtění. I když nebyl identifikován příčinný faktor v podobě jediného genu, budou pro klasifikaci haplotypů spojených s HB účinné úzce propojené markery, jako jsou jednonukleotidové polymorfismy, protože vazebná nerovnováha kultivované rýže se odhaduje na 100-200 kb, ačkoli u planých druhů (jako je např. rufipogon) může dosahovat až několika desítek kb (Mather et al., 2007; McNally et al., 2009; Huang et al., 2010). Tyto informace umožní navrhnout účinnější a efektivnější kombinaci křížení.
Segregantní analýza s následným sekvenováním nové generace může být užitečná pro mapování lokusů spojených s HB, stejně jako konvenční mapování vazeb (např. analýza lokusů kvantitativních znaků), protože HB segreganty často vykazují odlišné znaky (týkající se znaků, jako je výška rostlin, počet palic a plodnost) od normálních růstových segregantů v každé populaci křížení. Důležitou roli při fenotypizaci v laboratoři a na rýžovém poli může hrát také získávání obrazových dat pomocí digitální kamery a nakládání dronů. Toto úsilí poskytne cenné informace nejen šlechtitelům rýže, ale také evolučním biologům.
Pokud je navíc identifikována alela spojená s HB, může přerušení genů pomocí iontového svazku nebo editace genomu pomoci překonat HB, což je příkladem hybridní sterility F1 u rýže (Koide et al., 2018; Xie et al., 2019).
Prohlášení o dostupnosti dat
Všechny soubory dat prezentované v této studii jsou součástí článku/přílohového materiálu.
Příspěvek autora
Autor potvrzuje, že je jediným autorem této práce a schválil její zveřejnění.
Střet zájmů
Autoři prohlašují, že výzkum byl prováděn bez jakýchkoli komerčních nebo finančních vztahů, které by mohly být chápány jako potenciální střet zájmů.
Poděkování
Rád bych tento článek věnoval památce Dr. Darshana Brara. He was an eminent rice breeder who vigorusly introduced useful genes carried by wild species into cultivars beyond reproductive barriers.
Allard, R. W. (1960). Principy šlechtění rostlin. New York, NY: John Wily and Sons, Inc.
Google Scholar
Dobzhansky, T. (1970). Genetika evolučního procesu. New York, NY: Columbia University Press.
Google Scholar
Grant, V. (1971). Specifikace rostlin. New York, NY: Columbia University Press.
Google Scholar
Ikehashi, H., and Fujimaki, H. (1980). „Modified bulk population method for rice breeding,“ in Proceedings of the International Rice Research Conference on Innovative approaches to rice breeding, (Los Baños, PA: International Rice Research Institute), 163-182.
Google Scholar
Oka, H.-I. (1988). Původ pěstované rýže. Tokyo: JSSP.
Google Scholar
.