- Introduktion
- Den genetiska grunden för hybriduppdelning i ris
- Molekylära mekanismer som ligger till grund för hybriddestruktion
- Hybriduppdelning i konventionell risförädling
- Hybridförlust i genomikassisterad förädling
- Diskussion
- Data Availability Statement
- Author Contributions
- Intressekonflikt
- Acknowledgments
Introduktion
Förädlare och forskare har varit intresserade av hybridisering och introgression mellan divergerande genotyper som den potentiella drivkraften som leder till ekologisk divergens hos avkomman (Rieseberg et al., 2007; Arnold et al., 2012). Omvänt kan hybridisering ibland medföra en minskad hybridfödlighet, såsom svaghet och/eller sterilitet i F1-generationen och senare generationer, även i fall av hybridisering mellan medlemmar av samma art.
Reducerad hybridfödlighet och/eller fertilitet som segregeras i F2-generationen eller senare generationer benämns hybridförstöring (HB), där recessiva alleler med nödvändighet är förknippade. Detta reproduktionshinder har observerats under lång tid hos både växter och djur (Dobzhansky, 1970; Grant, 1971). Många av de genetiska analyserna av detta hinder har visat att det åstadkoms genom en komplementär effekt mellan och/eller bland loci med differentierade alleler, vanligen kallad Bateson-Dobzhansky-Muller (BDM)-inkompatibilitet (Rieseberg och Willis, 2007). HB inbegriper nödvändigtvis inneboende postzygotiska reproduktionsbarriärer, t.ex. hybridinviabilitet (inklusive svaghet, nekros och kloros) och hybridsterilitet (hos hane, hona eller båda könscellerna); därför kan vissa forskare inte skilja HB från inviabilitet och sterilitet i F1-avkomman. Trots detta anser jag att HB är en lämplig klassificering av reproduktionshinder eftersom den antyder deras underliggande genetiska grund (dvs. inblandning av recessiva alleler).
Nyligen genomförda studier med hjälp av modellväxtarten Arabidopsis har gett en bättre förståelse för HB när det gäller dess molekylära mekanismer (Vaid och Laitinen, 2019). Omvänt, även om ris (Oryza species) är en modellgröda, är den nuvarande förståelsen av den genetiska grunden för HB fortfarande begränsad i denna art. Detta beror troligen på att HB inte är ett reproduktivt hinder i F1-hybrider, där man förväntar sig en högre spannmålsproduktion jämfört med de föräldrade inavlade linjerna, och troligen på att undermåliga plantor som segregerar i F2-generationer och senare generationer lätt kan väljas ut baserat på fenotypen från avelspopulationen. Svårigheten med genetisk kartläggning som orsakas av recessiv nedärvning kan också ligga bakom denna begränsning.
Här beskriver jag HB i ris korsningar samtidigt som jag hänvisar till information från Arabidopsis-studier, och diskuterar hur HB hanteras i risförädling.
Den genetiska grunden för hybriduppdelning i ris
Det anses att odlat ris, O. sativa japonica och indica, bildar ett artkomplex med sin förmodade stamfader (O. rufipogon) och vilda arter; dock observeras flera reproduktiva barriärer, inklusive HB, i korsningar mellan dem (Oka, 1988; Vaughan et al, 2003).
Såvitt jag vet var Dr. Oka den förste som beskrev den genetiska grunden för HB i intersubspecifika korsningar mellan japonica och indica, vilket förklarades av två komplementära gener (Oka, 1957). De ansvariga generna kartlades dock inte på kromosomer, eftersom molekylära markörer inte fanns tillgängliga vid den tiden.
I rishybrider har HB ofta beskrivits i intersubspecifika (O. sativa ssp. japonica × ssp. indica) och interspecifika (O. sativa × O. nivara och O. sativa × O. glumaepatula) korsningar (Oka, 1957; Sato och Morishima, 1988; Wu et al, 1995; Li et al., 1997; Fukuoka et al., 1998, 2005; Kubo och Yoshimura, 2002, 2005; Matsubara et al., 2007a,b, 2015; Yamamoto et al., 2007; Ichitani et al., 2012; för intersubspecifika korsningar; Sobrizal et al., 2001; Miura et al., 2008 för interspecifika korsningar) (tabell 1). De flesta av dessa HB-fall orsakades av BDM-inkompatibilitet på två loci och recessiva alleler, varvid 1/16 av F2-avkomman som var homozygot för recessiva alleler på båda loci uppvisade HB-fenotypen; dock uppvisade 4/16 av avkomman som var heterozygot på endast ett locus HB-fenotypen, beroende på korsningskombinationen (tabell 1 och figur 1A). I backcrosshybriderna var en locus redan fixerad med allelerna från den återkommande föräldern; därför uppvisade 1/4 av BCnF2-avkomman HB-fenotypen (tabell 1 och figur 1B).

Tabell 1. Genetisk grund för den hybriduppdelning som rapporterats i ris korsningar.
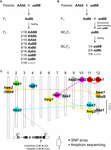
Figur 1. Den genetiska grunden för hybriduppdelning (HB). (A) Schematisk representation av HB-genotypen som segregeras i F2-avkomman. När föräldrarnas genotyper är AAbb och aaBB uppvisar 1/16 av F2-avkomman utan dominant allel, som anges med grå tecken (dvs. aabb), minskad livsduglighet och/eller fertilitet, även om de andra genotyperna är normala. I vissa ris korsningar uppvisar F2-avkomman med endast en dominant allel (dvs. Aabb, aaBb) också minskad livsduglighet och/eller fertilitet. (B) Schematisk representation av HB-genotypen som segregeras i BCnF2-avkomman. Dessutom uppvisar 1/4 av BCnF2-avkomman minskad livskraft och/eller fertilitet. (C) Kromosomal placering av de gener som ligger till grund för HB i ris korsningar. Generna är grovt kartlagda baserat på resultaten av genkartläggningen i varje studie. En uppsättning komplementära gener är sammanlänkade med den streckade linjen. Komplementära gener med hbd1 och hwf1 har inte rapporterats. Exemplet på den funktionella nukleotidpolymorfism som orsakar HB som publicerats av Yamamoto et al. (2010) visas, och denna information kommer att göra det möjligt för oss att genomföra en undersökning med hjälp av SNP-array eller amplikonsekvensering.
HBH-fenotypen hos rishybrider varierar beroende på korsningskombinationen. Till exempel visar genotypen hbd2/hbd2 hbd3/hbd3 svaghet, men ingen uppenbar frösterilitet. Genotypen hbd4/hbd4 hbd5/hbd5 uppvisar dock både svaghet och frösterilitet (Matsubara et al., 2007a, 2015; Yamamoto et al., 2007; tabell 1). Det finns också komplicerade fall där HB som visar både svaghet och frösterilitet observeras (genotyp hwe1/hwe1 hwe2/hwe2) eller HB som endast visar frösterilitet (hsa1/hsa1 hsa2/hsa2 hsa3/hsa3), trots samma korsningskombination, dvs. Asominori × IR24-korsning (Kubo och Yoshimura, 2002, 2005; tabell 1). Eftersom dessa HB har oberoende genetiska baser skulle minst 5/64 av F2-avkomman visa HB-fenotypen i denna korsning.
Hittills har flera uppsättningar av loci som är ansvariga för HB kartlagts till särskilda genomiska regioner med hjälp av DNA-markörer i korsningar mellan japonica- och indica-risvarianter (Chrs 7 och 10, Fukuoka et al, 1998; Chrs 1 och 12, Wu et al., 1995; Kubo och Yoshimura, 2002; Matsubara et al., 2015; Chrs 6 och 11, Fukuoka et al., 2005; Chrs 8, 9 och 12, Kubo och Yoshimura, 2005; Chrs 2 och 11, Matsubara et al., 2007a; Yamamoto et al., 2007, 2010; Chrs 11 och 12, Ichitani et al., 2012; tabell 1 och figur 1C). Resultaten av dessa studier visade att loci som ligger till grund för ris HB är gemensamma i vissa korsningar, men skiljer sig åt i andra korsningar, vilket tyder på att de är diversifierade i risgenomerna. I vissa fall delar till exempel den HB-allel som bärs av O. nivara, en art som är nära besläktad med incica-sorter, samma locus med en indica-sort, men den HB-allel som bärs av O. glumaepatula, en art som är nära besläktad med odlade sorter, delar inte locus med någon annan sort (tabell 1 och figur 1C). Det bör noteras att de HB-fall som beskrivs ovan orsakas av en uppsättning gener med stora effekter; det finns dock även fall som orsakas av en uppsättning gener med mindre effekter, t.ex. något minskad fröfruktbarhet, som inte kan förbises i risförädlingen, och som ännu inte har upptäckts.
Molekylära mekanismer som ligger till grund för hybriddestruktion
Autoimmun reaktion: Bomblies et al. (2007) rapporterade först att det autoimmuna svaret som en NB-LRR-sjukdomsresistensgen eller R-gen är förknippat med Arabidopsis HB i intraspecifika korsningar, även om detta autoimmuna svar huvudsakligen uttrycks som nekros i F1-plantor, och HB i F2-avkomman tycktes vara betingat av temperaturen. I Arabidopsis-hybrider har flera bevis för HB orsakad av det autoimmuna svaret beskrivits (Alcázar et al., 2009, 2010). Att NB-LRR-gener är involverade i HB tyder på att flera genomiska regioner kan förknippas med Arabidopsis HB; faktum är att den omfattande undersökning som Chae et al. (2014) genomförde stöder denna idé.
Också i rishybrider har HB som orsakats av ett autoimmunrespons rapporterats i en korsning av japonica × indica (Yamamoto et al., 2010). I detta fall uppstår HB i F2-plantor när genen hybrid breakdown 2 (hbd2), som kodar för kaseinkinas I och som bärs av indica-sorten, kombineras med ett kluster av R-gener som bärs av japonica-sorten. Eftersom dessa orsakande alleler sannolikt beter sig som delvis recessiva alleler kan genprodukterna vara inblandade (Matsubara et al., 2007a; Yamamoto et al., 2007, 2010).
Reciprok tystnad av duplicerade gener: Denna föreställning föreslogs först teoretiskt som en genetisk grund för hybridinkompatibilitet (Werth och Windham, 1991; Lynch och Force, 2000). I modellen förlorar en population funktionen på ett locus och behåller den på det andra, medan den andra populationen upplever den motsatta effekten. Följaktligen har 1/16 av F2-zygotterna inga funktionella gener (Lynch och Force, 2000). HB i Arabidopsis-hybrider som beskrivs av Bikard et al. (2009) och Vlad et al. (2010) är ett bra exempel på detta fenomen. I det här scenariot har fall som involverar epigenetisk tystnad också rapporterats (Agorio et al., 2017; Blevins et al., 2017).
Då ris har upplevt både helgenom- och segmentdubblering (Wang et al., 2007; Guo et al., 2019), kan ömsesidig tystnad vara förhärskande som en orsak till hybridinkompatibilitet. Hittills finns det dock inga bevis för att HB orsakas av ömsesidig tystnad av duplicerade gener i ris.
Oavhängigt av autoimmunresponsen och ömsesidig tystnad har en interaktion mellan ett DEAD-box RNA-helikas 18 och en annan gen (MORPHEUS MOLECULE 1 som en trolig kandidat) som orsakar HB i Arabidopsis-hybrider identifierats (Plötner et al., 2017; Vaid et al., 2020). Intressant nog minskar HB-fenotypen mellan F3- och F4-generationerna, vilket antyder epigenetisk reglering av genuttrycket.
I rishybrider, bland de tre loci för hybridsterilitet-a (hsa), visade Kubo et al. (2016) nyligen att hsa1-lokuset består av två gener, och att dessa gener kodar för ett DUF1618-protein respektive ett okarakteriserat protein med viss likhet med ett nukleotidbindande protein. De molekylära egenskaperna hos de återstående komplementära generna, hsa2 och hsa3, har inte rapporterats.
Hybriduppdelning i konventionell risförädling
I årtionden användes bulkpopulationsmetoden i stor utsträckning i konventionell risförädling. I denna metod odlas F4- eller F5-populationen efter korsning genom självbefruktning i bulk (i stället för genom nedkomstmetoden med ett enda frö) utan artificiellt urval, men den tidiga generationens population utsätts för naturligt urval och urval på grund av livskraft (Allard, 1960; Ikehashi och Fujimaki, 1980 för närmare information). I bulkpopulationsmetoden kan många svaga och/eller sterila genotyper förväntas elimineras från populationen innan en avancerad generationspopulation etableras; därför kan denna metod ge begränsad information om HB.
För introgression eller ackumulering av önskvärda egenskaper från donator(er) till en sort har risförädlare ofta utfört backcrossing eller multipel föräldrakorsning, följt av bulkpopulationsmetoden. Empiriskt har det länge varit känt att dessa korsningsmetoder gör det möjligt att mildra förlusten av urvalskandidater på grund av reproduktiva hinder, eftersom dessa metoder ofta minskar segregeringen av störande kombinationer av alleler som är förknippade med HB i en hybridavkomma. Mildringen av reproduktionsbarriärer genom dessa korsningsmetoder är dock oundvikligen beroende av föräldrarnas HB-genotyp.
Hybridförlust i genomikassisterad förädling
Publiceringen av referensgrödors genomsekvenser och utvecklingen av nästa generations sekvenseringstekniker har accelererat framstegen inom den molekylära förädlingen av grödor (Kole et al., 2015). I detta sammanhang har genomikassisterad förädling, såsom genomiskt urval baserat på genotyper av DNA-markörer för hela genomet, övervägts inom växtförädling (Spindel och Iwata, 2018).
I genomikassisterad förädling av självpollinerade grödor används ofta avancerad generationspopulationer, såsom rekombinanta inavelslinjer, som referenspopulationer, från vilka man erhåller genotyp- och fenotypdata för hela genomet. Därefter utförs urval baserat enbart på markörgenotyper i avkomman från tidiga generationer (t.ex. F2). I detta urvalssystem bör det noteras att referenspopulationen vanligtvis inte ger information om genomisk region för HB, eftersom svaga och/eller sterila avkommor borde ha eliminerats i de tidiga generationerna efter korsning. Om ett sådant urvalssystem antas måste vi därför överge några viktiga urvalskandidater. Alternativt kan vi välja oönskade kandidater så att HB blir uppenbart i senare generationer.
Diskussion
Trots det begränsade antalet studier i detta ämne verkar det som om de loci som ligger till grund för HB i ris korsningar är diversifierade snarare än gemensamma (tabell 1 och figur 1). Som beskrivits ovan bör information om fördelningen av HB-associerade alleler bland korsningsföräldrarna vara en förutsättning för risförädling, särskilt för att maximera effektiviteten av genomikassisterad förädling. Därför behövs ytterligare upptäckt av HB i ris korsningar och kartläggning av ansvariga gener i risgenomet. I Arabidopsis-hybrider har sådana uppgifter om R-gener och deras interagerande gener undersökts i stor utsträckning (Alcázar et al., 2010, 2014; Chae et al., 2014). Rapporteringen av hbd2-genen av Yamamoto et al. (2010) i en rishybrid är ett bra exempel på en omfattande undersökning som utförts med hjälp av SNP-arrayer eller amplikonsekvensering (figur 1C). Generna hbd3 och hsa1 är också kandidater för denna typ av undersökning. Så småningom kommer utvecklingen av kriterier som gör det möjligt att förutsäga HB baserat på föräldralinjernas genomiska information att fungera som ett viktigt verktyg för genomikassisterad förädling. Även om en orsakande faktor inte har identifierats som en enskild gen, kommer tätt sammankopplade markörer, t.ex. polymorfismer av enskilda nukleotider, att vara effektiva för att klassificera HB-associerade haplotyper, eftersom kopplingsdifferensen hos odlat ris har uppskattats till 100-200 kb, även om den hos vilda arter (t.ex. O. rufipogon) kan sträcka sig över flera tiotals kb (Mather et al., 2007; McNally et al., 2009; Huang et al., 2010). Denna information kommer att göra det möjligt att utforma en mer effektiv och ändamålsenlig korsningskombination.
Bulked segregant analysis followed by next-generation sequencing can be useful for the mapping of HB-associated loci, as well as conventional linkage mapping (such as quantitative trait locus analysis), because HB segregants often show distinguishable features (about traits such such as plant height, tiler number, and fertility) from normal growth segregants in each cross population. Förvärv av bilddata med hjälp av en digitalkamera och drönarladdning kan också spela en viktig roll vid fenotypning i laboratoriet och på risfältet. Dessa ansträngningar kommer att ge värdefull information inte bara till risförädlare utan även till evolutionsbiologer.
För övrigt, om en HB-associerad allel identifieras, kan genavbrott genom jonstrålning eller genomredigering hjälpa till att övervinna HB, vilket exemplifieras av F1-hybridsterilitet i ris (Koide et al., 2018; Xie et al, 2019).
Data Availability Statement
Alla dataset som presenteras i den här studien ingår i artikeln/det kompletterande materialet.
Author Contributions
Författaren bekräftar att han eller hon är den enda bidragsgivaren till det här arbetet och har godkänt det för publicering.
Intressekonflikt
Författarna förklarar att forskningen har utförts i avsaknad av kommersiella eller finansiella relationer som skulle kunna tolkas som en potentiell intressekonflikt.
Acknowledgments
Jag vill tillägna den här uppsatsen till minnet av Dr. Darshan Brar. Han var en framstående risförädlare som kraftfullt införde användbara gener som bärs upp av vilda arter i odlingar bortom reproduktiva hinder.
Allard, R. W. (1960). Principles of Plant Breeding (Principer för växtförädling). New York, NY: John Wily and Sons, Inc.
Google Scholar
Dobzhansky, T. (1970). Genetik för den evolutionära processen. New York, NY: Columbia University Press.
Google Scholar
Grant, V. (1971). Plant Speciation. New York, NY: Columbia University Press.
Google Scholar
Ikehashi, H., och Fujimaki, H. (1980). ”Modified bulk population method for rice breeding”, i Proceedings of the International Rice Research Conference on Innovative approaches to rice breeding, (Los Baños, PA: International Rice Research Institute), 163-182.
Google Scholar
Oka, H.-I. (1988). Det odlade risets ursprung. Tokyo: JSSP.
Google Scholar
.