Calea de apoptoză a perforinei/granzimei este principala cale de semnalizare utilizată de limfocitele citotoxice pentru a elimina celulele infectate și/sau transformate de virusuri. Studiile efectuate pe șoareci cu întreruperi genetice indică faptul că perforina, în combinație cu granzima, ar putea induce apoptoza. Este vitală pentru funcția efectoare citotoxică și are un rol indispensabil, dar nedefinit, în apoptoza mediată de granzime.
Perforina și granzima
Perforina este o proteină formatoare de pori și este cunoscută și sub denumirea de toxine cu granule citoplasmatice. Granzima este o familie de serin-proteaze înrudite structural, stocate în granulele citotoxice ale limfocitelor citotoxice (CL). Perforina și granzima induc în mod cooperativ apoptoza celulelor-țintă (figura 1). Granzima este necesară pentru declanșarea apoptozei celulelor-țintă, dar acestea depind de faptul că sunt livrate în mod corespunzător de către perforină. Atât perforina, cât și granzima se leagă la suprafața celulei-țintă ca parte a unui singur complex macromolecular asociat cu serglicina, ceea ce diminuează și mai mult probabilitatea de difuzie pasivă a granzimei.
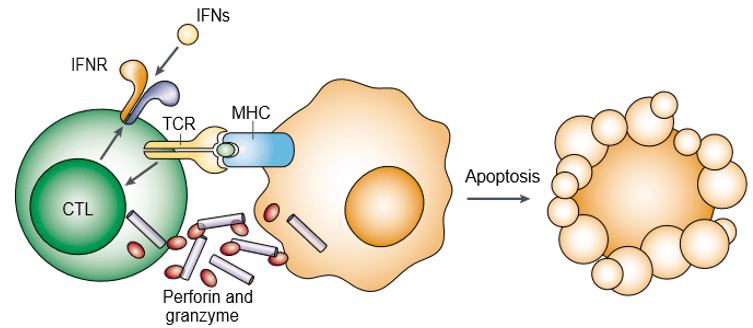
Figura 1. Perforina și granzima induc cooperativ apoptoza celulelor-țintă.
La om, există granzimele A, B, H, K și M, în timp ce la șoareci există granzimele A, B, C, D, E, F, G, K, L, M și N. Granzima A (GrA) și granzima B (GrB) sunt cele mai abundente granzime și au fost cele mai studiate. Funcțiile granzimelor A și B în inducerea apoptozei celulelor-țintă au fost studiate pe larg in vitro și sunt mai bine înțelese decât rolul perforinei la nivel molecular.
Procesul și reglarea căii de apoptoză Perforin/Granzimei
După ce sunt secretate de limfocitele citotoxice, granzimele pătrund în celulele-țintă, ceea ce reprezintă o etapă vitală în moartea celulară. Principalele proteine litice împachetate în granule sunt diferite granzime și proteina perforină formatoare de pori, care facilitează internalizarea granzimelor de către celule.
Granzima B declanșează în principal activarea caspazelor în mod indirect, mai degrabă decât prin procesarea directă a caspazelor. Ea realizează acest lucru prin activarea directă a membrilor proapoptotici „BH3-only” ai familiei BCL-2, cum ar fi agonistul morții cu domeniu de interacțiune cu BH3 (Bid). Bid, împreună cu proteinele proapoptotice Bax și/sau Bak din familia BCL-2, duce la scurgerea în citosol a mediatorilor mitocondriali proapoptotici, cum ar fi citocromul c. Eliberarea de citocrom c activează pro caspaza-9 și, prin legarea la factorul 1 de activare a proteazei apoptotice (Apaf-1), pro caspaza-9 devine caspaza-9 matură, care continuă să formeze apoptosomul și activează caspaza-3 din aval. Caspaza-3 activată este capabilă să cliveze substraturi specifice, cum ar fi ICAD (inhibitor al DNazei activate de caspază, CAD), permițând CAD să se transloce în nucleu pentru a fragmenta ADN-ul. Pe lângă Bid, granzima B poate inactiva Mcl-1, care este un membru al familiei antiapoptotice Bcl-2, pentru a elibera proteina pro-antiapoptotică Bim din familia Bcl-2 pe membrana mitocondrială externă. Iar granzima B poate, de asemenea, să medieze caspaza efectoare-3 și caspaza inițiatoare-8 pentru a regla calea apoptozei.
În contrast cu granzima B, granzima M nu depinde de mitocondriale pentru a le regla, dar granzima M poate activa granzima B prin scindarea inhibitorului de protează 9 (PI-9) care este inhibitorul granzimei B. Granzima M poate, de asemenea, să cliveze direct ICAD pentru a dezlănțui CAD ca și granzima B.
Granzima A induce pierderea potențialului membranei interne mitocondriale și eliberarea de specii reactive de oxigen (ROS). Ea generează nicks de ADN monocatenar, mai degrabă decât fragmente de ADN oligonucleosomal. Ca răspuns la ROS, complexul SET asociat ER, care include SET, Ape1, pp32, HMG2, NM23-H1 și TREX1, se translocă în nucleu, unde granzima A scindează trei membri ai complexului SET care sunt implicați în repararea ADN-ului: HMG2, Ape1 și SET.
Calea de apoptoză a perforinei/granzimei în boli
Studiile efectuate pe șoareci cu întreruperi genetice indică în mod clar că calea perforinei este implicată în mod intim în apărarea împotriva agenților patogeni virali și a celulelor transformate. Dar, au fost, de asemenea, demonstrate in vivo roluri nesuspectate pentru perforină ca mediator crucial al supravegherii imune a celulelor transformate spontan și în reglarea homeostaziei celulelor B și a memoriei celulelor T. În mod paradoxal, perforina joacă un rol esențial în patogeneza anumitor modele de boli autoimune, cum ar fi diabetul insulino-dependent la șoarecii diabetici non-obezi (NOD), dar ar putea suprima alte boli autoimune, inclusiv encefalomielita autoimună experimentală (EAE).
Se determină căile de apoptoză ale perforinei/granulei care reglează polarizarea și exocitoza granulelor, iar aceste căi ar putea fi perturbate de interacțiunea celulelor canceroase cu unele limfocite citotoxice. Descoperirile recente că intrarea granzimei în celulele țintă implică legarea la receptori membranari specifici și că unele tumori sintetizează inhibitori de serin protează care pot neutraliza granzimele au indicat noi căi potențiale prin care tumorile ar putea scăpa de atacul imunitar.
.