Evaluare | Biopsihologie | Comparată |Cognitivă | Dezvoltare | Limbaj | Diferențe individuale |Personalitate | Filosofie | Socială |
Metode | Statistică |Clinică | Educațională | Industrială |Profesională | Psihologie mondială |
Biologică:Genetică comportamentală – Psihologie evolutivă – Neuroanatomie – Neurochimie – Neuroendocrinologie – Neuroștiințe – Psihoneuroimunologie – Psihologie fiziologică – Psihofarmacologie(Index, Schiță)
| Creier: Hipotalamusul | ||
|---|---|---|
 |
||
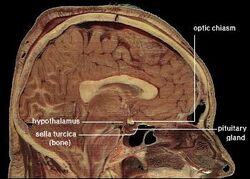
| Localizarea hipotalamusului uman | ||
 |
||
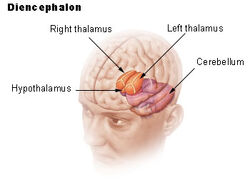
| Diencephalon | ||
| Latin | hypothalamus | |
| Gray’s | subiect #189 812 | |
| Parte de | ||
| Componente | ||
| Arteră | ||
| Arteră | ||
| Veină | ||
| BrainInfo/UW | hier-358 | |
| MeSH | A08.186.211.730.385.357 | |
Ipotalamusul face legătura între sistemul nervos și sistemul endocrin prin intermediul glandei pituitare (hipofiză). Hipotalamusul, (din greacă ὑποθαλαμος = sub talamus) este situat sub talamus, chiar deasupra trunchiului cerebral. Această glandă ocupă cea mai mare parte a regiunii ventrale a diencefalului. Se găsește în toate creierele mamiferelor. La om, are aproximativ dimensiunea unei migdale.
Ipotalamusul reglează anumite procese metabolice și alte activități ale sistemului nervos autonom. El sintetizează și secretă neurohormoni, adesea numiți hormoni de eliberare hipotalamică, iar aceștia, la rândul lor, stimulează sau inhibă secreția de hormoni hipofizari.
Ipotalamusul controlează temperatura corpului, foamea, setea, oboseala, furia și ciclurile circadiene.
- Intrări
- Stimuli olfactivi
- Stimuli transmiși prin sânge
- Steroizi
- Intrări neuronale
- Nuclee
- Ieșiri
- Proiecții neuronale
- Hormoni endocrini
- Controlul aportului alimentar
- Dimorfism sexual
- Răspunsurile la steroizii ovarieni
- Steroizi gonadici în viața neonatală a șobolanilor
- Androgenii la primate
- Alte influențe asupra dezvoltării hipotalamice
- Efectele îmbătrânirii asupra hipotalamusului
- Vezi și
- Imagini suplimentare
- Referințe & Bibliografie
- Lucrări
- Material suplimentar
- Lucrări
Intrări
Ipotalamusul este o regiune foarte complexă din creierul uman și chiar și nucleii mici din cadrul hipotalamusului sunt implicați în multe funcții diferite. Nucleul paraventricular, de exemplu, conține neuroni de oxitocină și vasopresină (numită și hormon antidiuretic) care se proiectează către hipofiza posterioară, dar conține și neuroni care reglează secreția de ACTH și TSH (care se proiectează către hipofiza anterioară), reflexele gastrice, comportamentul matern, tensiunea arterială, alimentația, răspunsurile imune și temperatura.
Ipotalamusul coordonează multe ritmuri circadiene hormonale și comportamentale, modele complexe de ieșiri neuroendocrine, mecanisme homeostatice complexe și multe comportamente importante. Prin urmare, hipotalamusul trebuie să răspundă la multe semnale diferite, dintre care unele sunt generate din exterior și altele din interior. Prin urmare, este bogat conectat cu multe părți ale SNC, inclusiv cu formațiunea reticulară a trunchiului cerebral și cu zonele autonome, cu creierul anterior limbic (în special amigdala, septul, banda diagonală a lui Broca și bulbii olfactivi, precum și cu cortexul cerebral).
Ipotalamusul este sensibil la:
- Lumină: lungimea zilei și fotoperioada pentru reglarea ritmurilor circadiene și sezoniere
- Stimuli olfactivi, inclusiv feromoni
- Steroizi, inclusiv steroizi gonadici și corticosteroizi
- Informații transmise pe cale neuronală care provin în special de la inimă, stomac, și tractul reproducător
- Intrare autonomă
- Stimuli sanguini, inclusiv leptină, grelină, angiotensină, insulină, hormoni hipofizari, citokine, concentrații plasmatice de glucoză și osmolaritate etc
- Stres
- Invadarea microorganismelor prin creșterea temperaturii corpului, resetarea în sus a termostatului organismului.
Stimuli olfactivi
Stimulii olfactivi sunt importanți pentru reproducere și funcția neuroendocrină la multe specii. De exemplu, dacă un șoarece gestant este expus la urina unui mascul „ciudat” în timpul unei perioade critice după coitus, atunci sarcina eșuează (efectul Bruce). Astfel, în timpul coitusului, o femelă de șoarece își formează o „memorie olfactivă” precisă a partenerului său, care persistă timp de mai multe zile.Indicii feromonali ajută la sincronizarea estrului la multe specii; la femei, sincronizarea menstruației poate apărea, de asemenea, ca urmare a indicilor feromonali, deși rolul feromonilor la om este pus la îndoială de unii.
Stimuli transmiși prin sânge
Hormonii peptidici au influențe importante asupra hipotalamusului și, pentru a face acest lucru, trebuie să eludeze bariera hemato-encefalică. Hipotalamusul este delimitat în parte de regiuni cerebrale specializate care nu au o barieră hemato-encefalică eficientă; endoteliul capilar din aceste locuri este fenestrat pentru a permite trecerea liberă chiar și a proteinelor mari și a altor molecule. Unele dintre aceste locuri sunt locurile de neurosecreție – neurohipofiza și eminența mediană. Cu toate acestea, altele sunt locurile în care creierul prelevează compoziția sângelui. Două dintre aceste situsuri, organul subfornical și OVLT (organum vasculosum of the lamina terminalis) sunt așa-numitele organe circumventriculare, unde neuronii sunt în contact intim atât cu sângele, cât și cu LCR. Aceste structuri sunt dens vascularizate și conțin neuroni osmoreceptivi și receptori de sodiu care controlează băutul, eliberarea de vasopresină, excreția de sodiu și apetitul pentru sodiu. Acestea conțin, de asemenea, neuroni cu receptori pentru angiotensină, factor natriuretic atrial, endotelină și relaxină, fiecare dintre aceștia fiind important în reglarea echilibrului hidric și electrolitic. Neuronii din OVLT și SFO se proiectează către nucleul supraoptic și nucleul paraventricular, precum și către zonele hipotalamice preoptice. Organele circumventriculare pot fi, de asemenea, locul de acțiune al interleukinelor pentru a declanșa atât febra, cât și secreția de ACTH, prin efecte asupra neuronilor paraventriculari.
Nu este clar cum obțin accesul necesar toate peptidele care influențează activitatea hipotalamică. În cazul prolactinei și leptinei, există dovezi de absorbție activă la nivelul plexului coroidian din sânge în LCR. Unii hormoni hipofizari au o influență negativă de feedback asupra secreției hipotalamice; de exemplu, hormonul de creștere se retroalimentează la nivelul hipotalamusului, dar modul în care acesta intră în creier nu este clar. Există, de asemenea, dovezi pentru acțiuni centrale ale prolactinei și TSH.
Steroizi
Ipotalamusul conține neuroni care sunt sensibili la steroizii gonadici și glucocorticoizi – (hormonii steroizi ai glandei suprarenale, eliberați ca răspuns la ACTH). De asemenea, conține neuroni specializați sensibili la glucoză (în nucleul arcuat și hipotalamusul ventromedial), care sunt importanți pentru apetit. Zona preoptică conține neuroni termosensibili; aceștia sunt importanți pentru secreția de TRH.
Intrări neuronale
Ipotalamusul primește multe intrări de la trunchiul cerebral; în special de la nucleul tractului solitar, de la locus coeruleus și de la măduva ventrolaterală. Secreția de oxitocină ca răspuns la supt sau la stimularea vagino-cervicală este mediată de unele dintre aceste căi; secreția de vasopresină ca răspuns la stimuli cardiovasculari proveniți de la chemoreceptorii din sinusul carotidian și din arcul aortic și de la receptorii volumului atrial de joasă presiune, este mediată de altele. La șobolan, stimularea vaginului determină, de asemenea, secreția de prolactină, iar acest lucru are ca rezultat o pseudo-graviditate în urma unei împerecheri infertile. La iepure, coitul provoacă ovulația reflexă. La oaie, stimularea cervicală în prezența unor niveluri ridicate de estrogen poate induce un comportament matern la o oaie virgină. Toate aceste efecte sunt mediate de hipotalamus, iar informația este transportată în principal prin căile spinale care se transmit în trunchiul cerebral. Stimularea mameloanelor stimulează eliberarea de oxitocină și prolactină și suprimă eliberarea de LH și FSH.
Stimulii cardiovasculari sunt transportați de nervul vag, dar vagul transmite, de asemenea, o varietate de informații viscerale, inclusiv, de exemplu, semnale care decurg din distensia gastrică pentru a suprima hrănirea. Din nou, aceste informații ajung la hipotalamus prin intermediul releelor din trunchiul cerebral.
Nuclee
Nucleii hipotalamici includ următoarele:
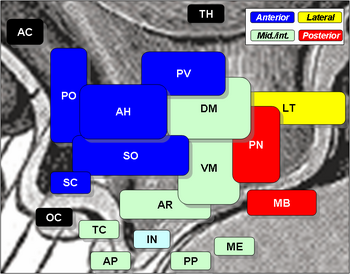
Nuclei hipotalamici
| Regiune | Arie | Nucleu | Nucleu | Funcție |
| Anterior | Medial | Nucleu preoptic medial |
|
|
| Supraoptic nucleu (SO) |
|
|||
| Nucleu paraventricular (PV) |
|
|||
| Nucleul hipotalamic anterior (AH) |
|
|||
| Nucleul suprachiasmatic (SC) |
|
|||
| Nucleul lateral | Nucleul preoptic lateral | |||
| Nucleul lateral (LT) |
|
|||
| Partea nucleului supraoptic (SO) |
|
|||
| Tuberal | Medial | Nucleul hipotalamic dorsomedial (DM) |
|
|
| Nucleul ventromedial (VM) |
. |
|||
Nucleul Arcuat (AR)
|
||||
| Lateral | . Nucleul lateral (LT) |
|
||
| Nucleul tuberal lateral | ||||
| Posterior | Medial | Nucleii mamilari (parte a corpilor mamilari) (MB) |
|
|
| Nucleul posterior (PN) |
|
|||
| Lateral | Nucleul lateral (LT) |
Vezi și: Consiliul lateral: nucleul preoptic ventrolateral
Ieșiri
Ieșirile hipotalamusului pot fi împărțite în două categorii: proiecții neuronale și hormoni endocrini.
Proiecții neuronale
Majoritatea sistemelor de fibre ale hipotalamusului se desfășoară în două direcții (bidirecțional).
- Proiecțiile către zonele caudale ale hipotalamusului trec prin fasciculul forebrain medial, tractul mamilotegmental și fasciculul longitudinal dorsal.
- Proiecțiile către zonele rostrale față de hipotalamus sunt purtate de tractul mamillo-talamic, de fornix și de stria terminală.
Hormoni endocrini
Hipotalamusul afectează sistemul endocrin și guvernează comportamentul emoțional, cum ar fi, furia și activitatea sexuală. Majoritatea hormonilor hipotalamici generați sunt distribuiți către hipofiză prin intermediul sistemului portal hipofizar. Hipotalamusul menține homeostazia acest lucru include o reglare a tensiunii arteriale, a ritmului cardiac și a temperaturii.
Hormonii hipotalamici primari sunt:
| Nume | Alte denumiri | Abbreviații | Localizare | Funcție | |
|---|---|---|---|---|---|
| Hormonul de eliberare a corticotropinei | Factorul de eliberare a corticotropinei, Corticoliberina | CRH, CRF | neuronii neuroendocrini parvocelulari din nucleul paraventricular | cu vasopresină, stimulează hipofiza anterioară să secrete ACTH | |
| Dopamina | hormonul inhibitor al prolactinei | DA, PIH | neuronii neuroendocrini din nucleul arcuat | inhibă secreția de prolactină din hipofiza anterioară | |
| Hormonul de eliberare a gonadotropinei | Hormonul de eliberare a hormonului de luteinizare | GnRH, LHRH | Neuronii neuroendocrini din nucleele preoptic medial și arcuat | stimulează hipofiza anterioară să secrete LH și FSH | |
| Hormonul de eliberare a hormonului de creștere | Factorul de eliberare a hormonului de creștere, somatocrinină | GHRH, GHRF, GRF | neuronii neuroendocrini ai nucleului arcuat | stimulează hipofiza anterioară să secrete hormonul de creștere | |
| Melatonina | Nucleul suprachiasmatic | ||||
| Somatostatina | Hormonul de creștere | .hormon inhibitor al creșterii, Factorul inhibitor al eliberării somatotropinei | SS, GHIH, SRIF | Neuronii neuroendocrini din nucleul periventricular | inhibă secreția hormonului de creștere din hipofiza anterioară |
| Hormonul de eliberare a tirotropinei | Factorul de eliberare a tirotropinei | Factorul de eliberare a tirotropinei, Tiroliberin, Protirelin | TRH, TRF | neuronii neuroendocrini parvocelulari din nucleele hipotalamice paraventriculare și anterioare | stimulează hipofiza anterioară să secrete TSH |
Vezi și: TSH: Hipocretină
Controlul aportului alimentar
Partea extremă laterală a nucleului ventromedial al hipotalamusului este responsabilă de controlul aportului alimentar. Stimularea acestei zone determină creșterea aportului alimentar. Lezarea bilaterală a acestei zone determină încetarea completă a aportului alimentar. Părțile mediale ale nucleului au un efect de control asupra părții laterale. Lezarea bilaterală a părții mediale a nucleului ventromedial determină hiperfagia și obezitatea animalului. Lezarea ulterioară a părții laterale a nucleului ventromedial la același animal produce încetarea completă a aportului alimentar.
Există diferite ipoteze legate de această reglare:
- Ipoteza lipostatică – această ipoteză susține că țesutul adipos produce un semnal umoral care este proporțional cu cantitatea de grăsime și acționează asupra hipotalamusului pentru a diminua aportul alimentar și a crește producția de energie. S-a evidențiat faptul că un hormon, leptina, acționează asupra hipotalamusului pentru a diminua aportul alimentar și a crește producția de energie.
- Ipoteza gastro-intestinală – hormoni gastrointestinali precum Grp, glucagoni, CCK și alții susțin că inhibă aportul alimentar. Alimentele care intră în tractul gastrointestinal declanșează eliberarea acestor hormoni care acționează asupra creierului pentru a produce sațietate. Creierul conține atât receptori CCK-A, cât și CCK-B.
- Ipoteza glucostatică – activitatea centrului de sațietate din nucleii ventromediali este probabil guvernată de utilizarea glucozei în neuroni. S-a emis ipoteza că atunci când utilizarea glucozei de către aceștia este scăzută și, în consecință, atunci când diferența de glucoză din sângele arterio-venos prin aceștia este scăzută, activitatea prin neuroni scade. În aceste condiții, activitatea centrului de hrănire este necontrolată, iar individul are senzația de foame. Ingestia de alimente este rapid crescută prin administrarea intraventriculară de 2-deoxiglucoză, scăzând, prin urmare, utilizarea glucozei în celule.
- Ipoteza termostatică – conform acestei ipoteze, o scădere a temperaturii corpului sub un anumit punct de referință stimulează apetitul, în timp ce o creștere peste punctul de referință inhibă apetitul.
Dimorfism sexual
Câțiva nuclei hipotalamici sunt dimorfici din punct de vedere sexual, adică există diferențe clare atât în structură, cât și în funcție între masculi și femele.
Câteva diferențe sunt evidente chiar și în neuroanatomia grosieră: cel mai notabil este nucleul dimorfic sexual din cadrul zonei preoptice, care este prezent numai la masculi. Cu toate acestea, majoritatea diferențelor sunt schimbări subtile în conectivitatea și sensibilitatea chimică a anumitor seturi de neuroni.
Importanța acestor schimbări poate fi recunoscută prin diferențele funcționale dintre masculi și femele. De exemplu, modelul de secreție a hormonului de creștere este dimorfic din punct de vedere sexual și acesta este unul dintre motivele pentru care, la multe specii, masculii adulți sunt mult mai mari decât femelele.
Răspunsurile la steroizii ovarieni
Alte dimorfisme funcționale izbitoare sunt în răspunsurile comportamentale la steroizii ovarieni ale adultului. Bărbații și femelele răspund diferit la steroizii ovarieni, în parte pentru că expresia neuronilor sensibili la estrogeni din hipotalamus este dimorfică din punct de vedere sexual, adică receptorii de estrogen sunt exprimați în diferite seturi de neuroni.
Estrogenul și progesteronul pot influența expresia genelor în anumiți neuroni sau pot induce modificări ale potențialului membranei celulare și activarea kinazelor, conducând la diverse funcții celulare non-genomice. Estrogenul și progesteronul se leagă la receptorii lor hormonali nucleari cognați, care se translocă în nucleul celular și interacționează cu regiuni ale ADN-ului cunoscute sub numele de elemente de răspuns hormonal (Hormone response elements – HRE) sau se leagă la situsul de legare al unui alt factor de transcripție. S-a demonstrat că receptorul estrogenic (ER) tranzactivează alți factori de transcripție în acest mod, în ciuda absenței unui element de răspuns estrogenic (ERE) în regiunea proximală a promotorului genei. ER și receptorii de progesteron (PR) sunt, în general, activatori de gene, cu o creștere a ARNm și o sinteză proteică ulterioară în urma expunerii la hormoni.
Creierul masculin și cel feminin diferă în ceea ce privește distribuția receptorilor de estrogen, iar această diferență este o consecință ireversibilă a expunerii neonatale la steroizi. Receptorii de estrogen (și receptorii de progesteron) se găsesc în principal în neuronii din hipotalamusul anterior și mediobazal, în special:
- zona preoptică (unde sunt localizați neuronii LHRH)
- nucleul periventricular (unde sunt localizați neuronii somatostatinei)
- hipotalamusul ventromedial (care este important pentru comportamentul sexual).
Steroizi gonadici în viața neonatală a șobolanilor
În viața neonatală, steroizii gonadici influențează dezvoltarea hipotalamusului neuroendocrin. De exemplu, aceștia determină capacitatea femelelor de a prezenta un ciclu reproductiv normal și a masculilor și femelelor de a prezenta comportamente reproductive adecvate în viața adultă.
- Dacă o femelă de șobolan este injectată o singură dată cu testosteron în primele câteva zile de viață postnatală (în timpul „perioadei critice” de influență a steroizilor sexuali), hipotalamusul este ireversibil masculinizat; șobolanul adult va fi incapabil să genereze o creștere a LH ca răspuns la estrogeni (o caracteristică a femelelor), dar va fi capabil să manifeste comportamente sexuale masculine (montarea unei femele receptive sexual).
- În schimb, un șobolan mascul castrat imediat după naștere va fi feminizat, iar adultul va prezenta comportamente sexuale feminine ca răspuns la estrogeni (receptivitate sexuală, lordose}.
Androgenii la primate
La primate, influența androgenilor asupra dezvoltării este mai puțin clară, iar consecințele sunt mai puțin complete. „Tomboyismul” la fete ar putea reflecta efectele androgenilor asupra creierului fetal, dar sexul de creștere în primii 2-3 ani este considerat de mulți ca fiind cel mai important factor determinant al identității de gen, deoarece în această fază fie estrogenul, fie testosteronul vor avea efecte permanente asupra creierului feminin sau masculin, influențând atât heterosexualitatea, cât și homosexualitatea.
Paradoxul este că efectele masculinizante ale testosteronului sunt mediate de estrogen. În interiorul creierului, testosteronul este aromatizat în (estradiol), care este principalul hormon activ pentru influențele de dezvoltare. Testiculul uman secretă niveluri ridicate de testosteron de la aproximativ săptămâna 8 de viață fetală până la 5-6 luni după naștere (o creștere perinatală similară a testosteronului este observată la multe specii), un proces care pare să stea la baza fenotipului masculin. Estrogenul din circulația maternă este relativ ineficient, parțial din cauza nivelurilor ridicate de proteine de legare a steroizilor aflate în circulație în timpul sarcinii.
Alte influențe asupra dezvoltării hipotalamice
Steroizii sexuali nu sunt singurele influențe importante asupra dezvoltării hipotalamice; în special, stresul pre-pubertal din primii ani de viață determină capacitatea hipotalamusului adult de a răspunde la un factor de stres acut. Spre deosebire de receptorii steroizilor gonadici, receptorii glucocorticoizi sunt foarte răspândiți în tot creierul; în nucleul paraventricular, aceștia mediază controlul prin feedback negativ al sintezei și secreției CRF, dar în alte părți rolul lor nu este bine înțeles.
Efectele îmbătrânirii asupra hipotalamusului
Studii la șoareci femele au arătat că atât nucleul supraoptic (SON), cât și nucleul paraventricular (PVN) pierd aproximativ o treime din celulele imunoreactive la IGF-1R odată cu îmbătrânirea normală. De asemenea, șoarecii bătrâni cu restricție calorică (CR) au pierdut un număr mai mare de celule neimunoreactive IGF-1R, menținând în același timp un număr similar de celule imunoreactive IGF-1R în comparație cu șoarecii bătrâni-Al. În consecință, șoarecii Old-CR prezintă un procent mai mare de celule IGF-1R imunoreactive, reflectând o sensibilitate hipotalamică crescută la IGF-1 în comparație cu șoarecii cu îmbătrânire normală.
Vezi și
- Axa HPA
- Sistemul hipofizar hipofizar hipofizar
- Sistemul hipofizar hipofizar suprarenal
- Leziuni ale hipotalamusului
- Factorul cerebral anterior mediu
- Neuroendocrinologie
- Zona preoptică
.
Imagini suplimentare
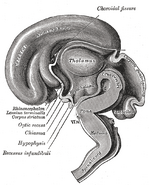
Adaugați o fotografie în această galerie
- http://www.cancer.gov/Templates/db_alpha.aspx?CdrID=46359
- http://www.sci.uidaho.edu/med532/hypothal.htm
- Diagrama nucleelor (psihopedagogie.org)
- Diagram of Nuclei (universe-review.ca)
- Diagram of Nuclei (utdallas.edu)
- Dacă nu se specifică altfel în tabel, atunci ref este: Guyton Ediția a 8-a
- Walter F., PhD. Borul. Fiziologie medicală: A Cellular And Molecular Approaoch, Elsevier/Saunders. Pag. 840
- http://thalamus.wustl.edu/course/hypoANS.html
- http://www.vivo.colostate.edu/hbooks/pathphys/endocrine/hypopit/overview.html
- Theologides A (1976). Metaboliți intermediari producători de anorexie. Am J Clin Nutr 29 (5): 552-8.
- John Money, „The concept of gender identity disorder in childhood and adolescence after 39 years”, Journal of Sex and Marital Therapy 20 (1994): 163-77.
- Romeo, Russell D, Rudy Bellani, Ilia N. Karatsoreos, Nara Chhua, Mary Vernov, Cheryl D. Conrad și Bruce S. McEwen (2005). Istoricul stresului și dezvoltarea pubertală interacționează pentru a modela plasticitatea axei hipotalamo-hipofizo-suprarenale. Endocrinology 147 (4): 1664-1674.
- Saeed O,Yaghmaie F,Garan SA,Gouw AM,Voelker MA,Sternberg H, Timiras PS. (2007). Celulele imunoreactive la receptorul receptorului factorului de creștere asemănător insulinei-1 sunt menținute selectiv în hipotalamusul paraventricular al șoarecilor restricționați caloric. Int J Dev Neurosci 25 (1): 23-8.
- Yaghmaie F, Saeed O, Garan SA, Voelker MA, Gouw AM, Freitag W, Sternberg H, Timiras PS (2006). Pierderea dependentă de vârstă a celulelor imunoreactive la receptorul receptorului factorului de creștere asemănător insulinei-1 în hipotalamusul supraoptic este redusă la șoarecii restricționați caloric. Int J Dev Neurosci 24 (7): 431-6.
- F. Yaghmaie, O. Saeed, S.A. Garan, A.M. Gouw, P. Jafar, J. Kaur, S. Nijjar, P.S. Timiras, H. Sternberg, M.A. Voelker (2007). Urmărirea modificărilor în sensibilitatea hipotalamică IGF-1 cu îmbătrânire și restricție calorică. Experimental Gerontology 42 (1-2): 148-149.
Referințe & Bibliografie
Lucrări
- Arees, E.A. și Mayer, B, (1967) Conexiuni anatomice între regiunile mediale și laterale ale hipotalamusului preocupate de aportul alimentar, Science 157: 1574-5
- Grossman, S.P. (1960) Mâncatul sau băutul provocat de stimularea directă adrenergică sau colinergică a hipotalamusului, Science 132: 301-2.
- Hetherington, A.W. și Ranson, S.W. (1942) Relația dintre diferite leziuni hipotalamice și adipozitatea la șobolan, 76: 475-99.
Material suplimentar
Lucrări
- Google Scholar
.
- BrainMaps at UCDavis Hypothalamus
- Sistem endocrin și hipotalamus
- High-Resolution Cytoarchitectural Primate Brain Atlases
- The Hypothalamus and Pituitary at endotexts.org
- Diagrama Nuclee (psycheducation.org)
- Diagrama Nuclee (universe-review.ca)
- Diagrama Nuclee (utdallas.edu)
|
v-d-e
Creierul uman: diencefalul (TA A14.1.08, GA 9.807) |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Epitalamus |
|
Suprafață
|
||||||||||||||
| Thalamus |
|
Suprafața
|
Materie cenușie/
nuclee |
|||||||||||||
| Hipotalamus |
|
|||||||||||||||
| Subtalamus |
Nucleul subtalamic – Zona incerta |
|||||||||||||||
| Sistemul ventricular: Ventriculul al treilea |
recesele: (recesiunea optică, recesiunea infundibulară, recesiunea suprapineală, recesiunea pineală) Solul hipotalamic – Tela corioidiană a celui de-al treilea ventricul Organul subforic Aperturi: Interventricular/Monro Comisura posterioară |
|||||||||||||||
|
v-d-e
Anatomia umană, sistemul endocrin: glande endocrine |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Axe hipotalamice/ hipofizare |
|
|
|
|
||||||||||||
| Glanda pineală |
Pinealocit – Corpora arenacea |
|||||||||||||||
| Isloți de pancreas |
Celula Alfa – Celula Beta – Celula Delta – Celula PP – Celula Epsilon |
|||||||||||||||
Creierul uman: Sistemul limbic
Amigdala – Girusul cingular – Girusul forțat – Hipocampul – Hipotalamusul – Corpul mamilar – Nucleus accumbens – Cortexul orbitofrontal – Girusul parahipocampal
.