- Introducere
- Bazele genetice ale ruperii hibrizilor la orez
- Mecanisme moleculare care stau la baza ruperii hibrizilor
- Hybrid Breakdown in Conventional Rice Breeding
- Pierderea hibrizilor în ameliorarea asistată de genomică
- Discuție
- Data Availability Statement
- Contribuții ale autorului
- Conflict de interese
- Recunoștințe
Introducere
Elevatorii și cercetătorii deopotrivă au fost interesați de hibridizarea și introgresia între genotipuri divergente ca potențial motor care conduce la divergența ecologică a progeniturilor (Rieseberg et al., 2007; Arnold et al., 2012). Dimpotrivă, hibridizarea poate implica uneori o fitness hibrid redusă, cum ar fi slăbiciunea și/sau sterilitatea în generațiile F1 și în generațiile ulterioare, chiar și în cazul hibridării între membrii aceleiași specii.
Viabilitatea și/sau fertilitatea hibridă redusă care segregă în generațiile F2 sau în generațiile ulterioare sunt denumite degradare hibridă (HB), în care sunt asociate în mod necesar alele recesive. Această barieră reproductivă a fost observată de mult timp atât la plante, cât și la animale (Dobzhansky, 1970; Grant, 1971). Multe dintre analizele genetice ale acestei bariere au arătat că ea se realizează printr-un efect complementar între și/sau între loci cu alele diferențiate, numit în mod obișnuit incompatibilitatea Bateson-Dobzhansky-Muller (BDM) (Rieseberg și Willis, 2007). HB implică în mod necesar bariere reproductive postzigotice intrinseci, cum ar fi inviabilitatea hibridă (inclusiv slăbiciune, necroză și cloroză) și sterilitatea hibridă (la nivelul gameților masculi, feminini sau ambii gameți); prin urmare, este posibil ca unii cercetători să nu facă distincția între HB și inviabilitatea și sterilitatea în progenitura F1. Cu toate acestea, cred că HB este o clasificare convenabilă pentru barierele de reproducere, deoarece implică baza genetică subiacentă a acestora (de exemplu, implicarea alelelor recesive).
Studii recente care utilizează specia de plante model Arabidopsis au oferit o mai bună înțelegere a HB în ceea ce privește mecanismele sale moleculare (Vaid și Laitinen, 2019). În schimb, deși orezul (specia Oryza) este o cultură model, înțelegerea actuală a bazei genetice a HB rămâne limitată la această specie, probabil pentru că HB nu reprezintă o barieră reproductivă în hibrizii F1, la care se așteaptă o producție mai mare de boabe în comparație cu liniile parentale consangvinizate, și probabil pentru că plantele inferioare care segregă în F2 și în generațiile ulterioare pot fi ușor selectate pe baza fenotipului din populația de reproducere. Dificultatea în cartografierea genetică cauzată de moștenirea recesivă poate fi, de asemenea, în spatele acestei limitări.
În cele ce urmează, prezint HB în încrucișările de orez, făcând referire la informațiile furnizate de studiile Arabidopsis, și discut despre modul în care HB este tratat în ameliorarea orezului.
Bazele genetice ale ruperii hibrizilor la orez
Se consideră că orezul cultivat, O. sativa japonica și indica, formează un complex de specii cu progenitorul lor putativ (O. rufipogon) și speciile sălbatice; cu toate acestea, în încrucișările dintre acestea se observă multiple bariere reproductive, inclusiv HB, (Oka, 1988; Vaughan et al, 2003).
Din câte știu eu, Dr. Oka a fost primul care a descris baza genetică a HB în încrucișările intersubspecifice între japonica și indica, care a fost explicată prin două gene complementare (Oka, 1957). Cu toate acestea, genele responsabile nu au fost cartografiate pe cromozomi, deoarece markerii moleculari nu erau disponibili la acea vreme.
În hibrizii de orez, HB a fost adesea descrisă în încrucișările intersubspecifice (O. sativa ssp. japonica × ssp. indica) și interspecifice (O. sativa × O. nivara și O. sativa × O. glumaepatula) (Oka, 1957; Sato și Morishima, 1988; Wu et al, 1995; Li et al., 1997; Fukuoka et al., 1998, 2005; Kubo și Yoshimura, 2002, 2005; Matsubara et al., 2007a,b, 2015; Yamamoto et al., 2007; Ichitani et al., 2012;pentru încrucișări intersubspecifice; Sobrizal et al., 2001; Miura et al., 2008 pentru încrucișări interspecifice) (tabelul 1) (tabelul 1). Cele mai multe dintre aceste cazuri de HB au fost cauzate de incompatibilitatea BDM cu două locații și de alele recesive, prin care 1/16 din descendența F2 care a fost homozigotă pentru alele recesive la ambii loci a prezentat fenotipul HB; cu toate acestea, 4/16 din descendența care a fost heterozigotă doar la un singur locus a prezentat fenotipul HB, în funcție de combinația de încrucișări (tabelul 1 și figura 1A). În hibrizii retrocrucișați, un locus a fost deja fixat cu alelele de la părintele recurent; prin urmare, 1/4 din progenitura BCnF2 a prezentat fenotipul HB (tabelul 1 și figura 1B).

Tabel 1. Baza genetică a descompunerii hibride raportate în încrucișările de orez.
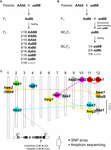
Figura 1. Baza genetică a descompunerii hibride (HB). (A) Reprezentare schematică a genotipului HB care segregă în progenitura F2. Atunci când genotipurile parentale sunt AAbb și aaBB, 1/16 din descendenții F2 fără alela dominantă, care este indicată prin caracterul gri (adică aabb), prezintă o viabilitate și/sau fertilitate redusă, deși celelalte genotipuri sunt normale. În unele încrucișări de orez, progeniturile F2 cu o singură alelă dominantă (de exemplu, Aabb, aaBb) prezintă, de asemenea, viabilitate și/sau fertilitate redusă. (B) Reprezentare schematică a genotipului HB care segregă în progeniturile BCnF2. În plus, 1/4 din progenitura BCnF2 prezintă viabilitate și/sau fertilitate redusă. (C) Localizarea cromozomială a genelor care stau la baza HB în încrucișările de orez. Genele sunt cartografiate aproximativ pe baza rezultatelor cartografierii genice din fiecare studiu. Un set de gene complementare sunt conectate prin linia punctată. Genele complementare cu hbd1 și hwf1 nu au fost raportate. Este prezentat exemplul polimorfismului funcțional de nucleotide care cauzează HB publicat de Yamamoto et al. (2010), iar aceste informații ne vor permite să realizăm un studiu prin matrice SNP sau secvențiere de ampliconi.
Fenotipul HB la hibrizii de orez variază în funcție de combinația de încrucișare. De exemplu, genotipul hbd2/hbd2 hbd3/hbd3 prezintă slăbiciune, dar nu prezintă o sterilitate evidentă a semințelor. Cu toate acestea, genotipul hbd4/hbd4 hbd5/hbd5 prezintă atât slăbiciune, cât și sterilitate a semințelor (Matsubara et al., 2007a, 2015; Yamamoto et al., 2007; Tabelul 1). Există, de asemenea, cazuri complicate în care se observă HB care prezintă atât slăbiciune, cât și sterilitate a semințelor (genotipul hwe1/hwe1 hwe2/hwe2) sau HB care prezintă doar sterilitate a semințelor (hsa1/hsa1 hsa2/hsa2 hsa3/hsa3), în ciuda aceleiași combinații de încrucișare, și anume, încrucișarea Asominori × IR24 (Kubo și Yoshimura, 2002, 2005; tabelul 1). Deoarece aceste HB-uri au baze genetice independente, cel puțin 5/64 din progeniturile F2 ar prezenta fenotipul HB în această încrucișare.
Până în prezent, mai multe seturi de loci responsabili pentru HB au fost cartografiate în anumite regiuni genomice folosind markeri ADN în încrucișările dintre soiurile de orez japonica și indica (Chrs 7 și 10, Fukuoka et al, 1998; Chrs 1 și 12, Wu et al., 1995; Kubo și Yoshimura, 2002; Matsubara et al., 2015; Chrs 6 și 11, Fukuoka et al., 2005; Chrs 8, 9 și 12, Kubo și Yoshimura, 2005; Chrs 2 și 11, Matsubara et al., 2007a; Yamamoto et al., 2007, 2010; Chrs 11 și 12, Ichitani et al., 2012; tabelul 1 și figura 1C). Rezultatele acestor studii au arătat că loci care stau la baza orezului HB sunt comune în unele încrucișări, dar diferă în alte încrucișări, ceea ce sugerează diversificarea lor în genomurile de orez. De exemplu, în unele cazuri, alela HB purtată de O. nivara, o specie înrudită îndeaproape cu soiurile incica, împarte același locus cu un soi indica; cu toate acestea, alela HB purtată de O. glumaepatula, o specie înrudită îndeaproape cu soiurile cultivate, nu împarte acest locus cu niciun alt soi (tabelul 1 și figura 1C). Trebuie remarcat faptul că cazurile HB descrise mai sus sunt cauzate de un set de gene cu efecte majore; cu toate acestea, există, de asemenea, cazuri cauzate de un set de gene cu efecte minore, de exemplu, fertilitatea ușor redusă a semințelor, care nu pot fi trecute cu vederea în ameliorarea orezului, care nu au fost încă detectate.
Mecanisme moleculare care stau la baza ruperii hibrizilor
Răspuns autoimun: Bomblies et al. (2007) au raportat pentru prima dată că răspunsul autoimun care o genă de rezistență la boli NB-LRR sau o genă R este asociat cu HB la Arabidopsis în încrucișările intraspecifice, deși acest răspuns autoimun este exprimat în principal sub formă de necroză în plantele F1, iar HB în progenitura F2 pare să fie condiționat de temperatură. La hibrizii de Arabidopsis, au fost descrise mai multe linii de dovezi de HB cauzate de răspunsul autoimun (Alcázar et al., 2009, 2010). Implicarea genelor NB-LRR în HB sugerează că mai multe regiuni genomice pot fi asociate cu HB la Arabidopsis; de fapt, studiul amplu realizat de Chae et al. (2014) susține această idee.
De asemenea, la hibrizii de orez, HB cauzată de un răspuns autoimun a fost raportată într-o încrucișare japonica × indica (Yamamoto et al., 2010). În acest caz, HB apare la plantele F2 atunci când gena hibrid breakdown 2 (hbd2), care codifică cazeina kinaza I și este purtată de soiul indica, este combinată cu un grup de gene R purtate de soiul japonica. Deoarece este probabil ca aceste alele cauzatoare să se comporte ca niște alele parțial recesive, produsele genice pot fi implicate (Matsubara et al., 2007a; Yamamoto et al., 2007, 2010).
Silențierea reciprocă a genelor duplicate: această noțiune a fost propusă pentru prima dată în mod teoretic ca o bază genetică pentru incompatibilitatea hibridă (Werth și Windham, 1991; Lynch și Force, 2000). În acest model, o populație pierde funcția la un locus și o păstrează la celălalt, în timp ce cealaltă populație experimentează efectul opus. În consecință, 1/16 din zigoții F2 nu au gene funcționale (Lynch și Force, 2000). HB la hibrizii de Arabidopsis descrisă de Bikard et al. (2009) și Vlad et al. (2010) este un bun exemplu al acestui fenomen. În acest scenariu, au fost raportate, de asemenea, cazuri care implică silențiozitatea epigenetică (Agorio et al., 2017; Blevins et al., 2017).
Pentru că orezul a cunoscut atât duplicarea întregului genom, cât și a segmentelor (Wang et al., 2007; Guo et al., 2019), silențiozitatea reciprocă ar putea fi prevalentă ca o cauză a incompatibilității hibride. Cu toate acestea, până în prezent, nu există nicio dovadă de HB cauzată de silențierea reciprocă a genelor duplicate la orez.
Independent de răspunsul autoimun și de silențierea reciprocă, a fost identificată o interacțiune între o ARN-elicază 18 DEAD-box RNA și o altă genă (MORPHEUS MOLECULE 1 ca un candidat probabil) care provoacă HB în hibrizii Arabidopsis (Plötner et al., 2017; Vaid et al., 2020). În mod interesant, fenotipul HB se reduce între generațiile F3 și F4, ceea ce implică o reglare epigenetică a expresiei genice.
La hibrizii de orez, printre cei trei loci de sterilitate hibridă-a (hsa), Kubo et al. (2016) au arătat recent că locusul hsa1 este format din două gene și că aceste gene codifică o proteină DUF1618 și, respectiv, o proteină necaracterizată cu o oarecare asemănare cu o proteină de legare a nucleotidelor. Caracteristicile moleculare ale celorlalte gene complementare, hsa2 și hsa3, nu au fost raportate.
Hybrid Breakdown in Conventional Rice Breeding
Pe parcursul a zeci de ani, metoda bulk-population a fost utilizată pe scară largă în ameliorarea convențională a orezului. În această metodă, după încrucișare, populația F4 sau F5 este crescută prin autofertilizare în masă (mai degrabă decât prin metoda descendenței cu o singură sămânță) fără selecție artificială, dar populația din prima generație este supusă selecției naturale și a viabilității (Allard, 1960; Ikehashi și Fujimaki, 1980 pentru detalii). În metoda de populare în masă, este de așteptat ca multe genotipuri slabe și/sau sterile să fie eliminate din populație înainte de stabilirea unei populații de generație avansată; prin urmare, această metodă poate furniza informații limitate despre HB.
Pentru introgresia sau acumularea trăsăturilor dezirabile de la donator(i) la un soi, amelioratorii de orez au efectuat adesea încrucișări inverse sau încrucișări parentale multiple, urmate de metoda de populare în masă. Din punct de vedere empiric, se știe de mult timp că aceste metode de încrucișare permit atenuarea pierderii candidaților la selecție prin bariere reproductive, deoarece aceste metode reduc adesea segregarea combinațiilor disruptive de alele asociate cu HB într-o descendență hibridă. Cu toate acestea, atenuarea barierelor de reproducere prin aceste metode de încrucișare depinde în mod inevitabil de genotipul HB al părinților.
Pierderea hibrizilor în ameliorarea asistată de genomică
Publicarea secvențelor genomului culturilor de referință și dezvoltarea tehnologiilor de secvențiere de generație următoare au accelerat progresul ameliorării moleculare a culturilor (Kole et al., 2015). În acest context, ameliorarea asistată de genomică, cum ar fi selecția genomică bazată pe genotipurile markerilor ADN la nivelul întregului genom, a fost luată în considerare în ameliorarea culturilor (Spindel și Iwata, 2018).
În ameliorarea asistată de genomică a culturilor autogame, populațiile de generație avansată, cum ar fi liniile consangvinizate recombinate, sunt adesea utilizate ca populații de referință, de la care se obțin date genotipice și fenotipice la nivelul întregului genom. Ulterior, selecția bazată numai pe genotipurile markerilor se efectuează în descendența generațiilor timpurii (de exemplu, F2). În această schemă de selecție, trebuie remarcat faptul că populația de referință nu furnizează, de obicei, informații despre regiunea genomică pentru HB, deoarece progeniturile slabe și/sau sterile ar fi trebuit să fie eliminate în primele generații după încrucișare. Prin urmare, dacă se adoptă o astfel de schemă de selecție, va trebui să renunțăm la unii candidați importanți pentru selecție. Alternativ, putem selecta candidați indezirabili, astfel încât HB să devină evident în generațiile ulterioare.
Discuție
În ciuda numărului limitat de studii pe această temă, se pare că loci care stau la baza HB în încrucișările de orez sunt mai degrabă diversificați decât comuni (tabelul 1 și figura 1). După cum s-a descris mai sus, informațiile privind distribuția alelelor asociate cu HB în rândul părinților încrucișați ar trebui să fie o condiție prealabilă pentru ameliorarea orezului, în special pentru a maximiza eficacitatea ameliorării asistate de genomică. Prin urmare, este necesară detectarea suplimentară a HB în încrucișările de orez și cartografierea genelor responsabile în genomul orezului. În hibrizii de Arabidopsis, astfel de date despre genele R și genele care interacționează cu acestea au fost cercetate pe larg (Alcázar et al., 2010, 2014; Chae et al., 2014). Raportarea genei hbd2 de către Yamamoto et al. (2010) la un hibrid de orez este un bun exemplu de studiu amplu realizat cu ajutorul matricelor SNP sau al secvențierii ampliconice (figura 1C). Genele hbd3 și hsa1 sunt, de asemenea, candidate pentru acest tip de studiu. În cele din urmă, dezvoltarea unor criterii care să ne permită să prezicem HB pe baza informațiilor genomice ale liniilor parentale va servi drept un instrument important pentru ameliorarea asistată de genomică. Chiar dacă nu a fost identificat un factor cauzal ca fiind o singură genă, markerii strâns legați, cum ar fi polimorfismele cu un singur nucleotid, vor fi eficienți pentru clasificarea haplotipurilor asociate cu HB, deoarece s-a estimat că dezechilibrul de legătură al orezului cultivat se extinde la 100-200 kb, deși cel al speciilor sălbatice (cum ar fi O. rufipogon) se poate extinde pe câteva zeci de kb (Mather et al., 2007; McNally et al., 2009; Huang et al., 2010). Aceste informații vor permite conceperea unei combinații încrucișate mai eficiente și mai eficace.
Analiza segregantelor în bloc urmată de secvențierea de generație următoare poate fi utilă pentru cartografierea lociilor asociați cu HB, precum și pentru cartografierea convențională a legăturilor (cum ar fi analiza locusurilor de trăsături cantitative), deoarece segregantele HB prezintă adesea caracteristici care se disting (cu privire la trăsături cum ar fi înălțimea plantelor, numărul de tuberculi și fertilitatea) de segregantele cu creștere normală din fiecare populație încrucișată. Achiziționarea de date imagistice cu ajutorul unei camere digitale și încărcarea dronelor poate juca, de asemenea, un rol important în fenotiparea în laborator și în câmpul de orezărie. Aceste eforturi vor furniza informații valoroase nu numai pentru amelioratorii de orez, ci și pentru biologii evolutivi.
În plus, dacă se identifică o alelă asociată cu HB, întreruperea genei prin fascicul de ioni sau editarea genomului poate ajuta la depășirea HB, așa cum este exemplificat de sterilitatea hibridă F1 la orez (Koide et al., 2018; Xie et al, 2019).
Data Availability Statement
Toate seturile de date prezentate în acest studiu sunt incluse în articol/materialul suplimentar.
Contribuții ale autorului
Autorul confirmă că este singurul contributor al acestei lucrări și a aprobat-o pentru publicare.
Conflict de interese
Autorii declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Recunoștințe
Dedic această lucrare memoriei Dr. Darshan Brar. El a fost un eminent ameliorator de orez care a introdus cu vigoare genele utile purtate de speciile sălbatice în cultivare dincolo de barierele reproductive.
Allard, R. W. (1960). Principii de ameliorare a plantelor. New York, NY: John Wily and Sons, Inc.
Google Scholar
Dobzhansky, T. (1970). Genetica procesului evolutiv. New York, NY: Columbia University Press.
Google Scholar
Grant, V. (1971). Speciația plantelor. New York, NY: Columbia University Press.
Google Scholar
Ikehashi, H., și Fujimaki, H. (1980). „Modified bulk population method for rice breeding,” în Proceedings of the International Rice Research Conference on Innovative approaches to rice breeding, (Los Baños, PA: International Rice Research Institute), 163-182.
Google Scholar
Oka, H.-I. (1988). Originea orezului cultivat. Tokyo: JSSP.
Google Scholar
.