Ocena | Biopsychologia | Porównawcza | Poznawcza | Rozwojowa | Językowa | Różnic indywidualnych | Osobowościowa | Filozoficzna | Społeczna |
Metody | Statystyczna | Kliniczna | Edukacyjna | Przemysłowa | Zawodowa | Psychologia Świata |
Biologiczna:Genetyka behawioralna – Psychologia ewolucyjna – Neuroanatomia – Neurochemia – Neuroendokrynologia -Neurobiologia – Psychoneuroimmunologia – Psychologia fizjologiczna – Psychofarmakologia(Indeks, Zarys)
| Mózg: Podwzgórze | ||
|---|---|---|
 |
||
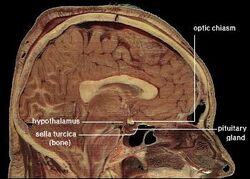
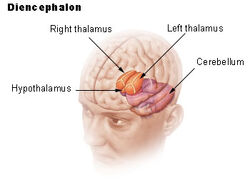
| Lokalizacja ludzkiego podwzgórza | ||
 |
||
| Dienchephalon | ||
| Łacina | hypothalamus | |
| Gray’s | subject #189 812 | |
| Part. of | ||
| Komponenty | ||
| Tętnica | ||
| Wein | ||
| BrainInfo/UW | hier-.358 | |
| MeSH | A08.186.211.730.385.357 | |
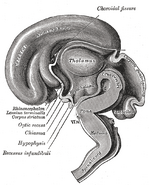
Podwzgórze łączy układ nerwowy z układem endokrynnym poprzez przysadkę mózgową (hypophysis). Podwzgórze, (z greckiego ὑποθαλαμος = pod wzgórzem) znajduje się poniżej wzgórza, tuż nad pniem mózgu. Gruczoł ten zajmuje większą część brzusznej części diencephalonu. Występuje w mózgach wszystkich ssaków. U ludzi jest on mniej więcej wielkości migdała.
Podwzgórze reguluje niektóre procesy metaboliczne i inne czynności Autonomicznego Układu Nerwowego. Syntetyzuje i wydziela neurohormony, często nazywane hormonami uwalniającymi podwzgórze, a te z kolei stymulują lub hamują wydzielanie hormonów przysadki mózgowej.
Podwzgórze kontroluje temperaturę ciała, głód, pragnienie, zmęczenie, gniew i cykle okołodobowe.
- Wejścia
- Bodźce węchowe
- Bodźce przenoszone przez krew
- Steroidy
- Wejścia neuronalne
- Jądra
- Wyjścia
- Projekcje neuronalne
- Hormony endokrynne
- Kontrola przyjmowania pokarmów
- Dymorfizm płciowy
- Odpowiedzi na sterydy jajnikowe
- Steroidy gonadalne w życiu noworodkowym szczurów
- Androgeny w naczelnych
- Inne wpływy na rozwój podwzgórza
- Wpływ starzenia się na podwzgórze
- Zobacz także
- Obrazy dodatkowe
- Referencje & Bibliografia
- Prace
- Materiały dodatkowe
- Papiery
Wejścia
Podwzgórze jest bardzo złożonym regionem w mózgu człowieka, a nawet małe jądra w podwzgórzu są zaangażowane w wiele różnych funkcji. Jądro przyśrodkowe na przykład zawiera neurony oksytocyny i wazopresyny (zwanej także hormonem antydiuretycznym), które rzutują do tylnej przysadki, ale także zawiera neurony, które regulują wydzielanie ACTH i TSH (które rzutują do przedniej przysadki), odruchy żołądkowe, zachowania macierzyńskie, ciśnienie krwi, karmienie, odpowiedzi immunologiczne i temperaturę.
Podwzgórze koordynuje wiele hormonalnych i behawioralnych rytmów okołodobowych, złożonych wzorów neuroendokrynnych wyjść, złożonych mechanizmów homeostatycznych i wielu ważnych zachowań. Podwzgórze musi zatem odpowiadać na wiele różnych sygnałów, z których niektóre są generowane zewnętrznie, a niektóre wewnętrznie. W ten sposób jest ono bogato połączone z wieloma częściami OUN, w tym z tworem siatkowatym pnia mózgu i strefami autonomicznymi, przedmózgowiem limbicznym (szczególnie z migdałkiem, przegrodą, ukośnym pasmem Broca i opuszkami węchowymi oraz korą mózgową).
Podwzgórze jest wrażliwe na:
- Światło: długość dnia i fotoperiod w celu regulacji rytmów okołodobowych i sezonowych
- Bodźce zapachowe, w tym feromony
- Steroidy, w tym steroidy gonadalne i kortykosteroidy
- Neuralnie przekazywane informacje powstające w szczególności z serca, żołądka, i układu rozrodczego
- Wejścia autonomiczne
- Bodźce pochodzące z krwi, w tym leptyna, grelina, angiotensyna, insulina, hormony przysadki mózgowej, cytokiny, stężenie glukozy w osoczu i osmolarność itp
- Stres
- Wprowadzanie drobnoustrojów poprzez podwyższenie temperatury ciała, przestawienie termostatu ciała w górę.
Bodźce węchowe
Bodźce węchowe są ważne dla reprodukcji i funkcji neuroendokrynnych u wielu gatunków. Na przykład, jeśli ciężarna mysz jest narażona na mocz „obcego” samca w krytycznym okresie po współżyciu, to ciąża się nie udaje (efekt Bruce’a). Tak więc podczas współżycia samica myszy tworzy precyzyjną „pamięć węchową” swojego partnera, która utrzymuje się przez kilka dni.Wskazówki feromonalne pomagają w synchronizacji rui u wielu gatunków; u kobiet zsynchronizowana menstruacja może również wynikać ze wskazówek feromonalnych, chociaż rola feromonów u ludzi jest przez niektórych kwestionowana.
Bodźce przenoszone przez krew
Hormony peptydowe mają ważny wpływ na podwzgórze, i aby to zrobić, muszą omijać barierę krew-mózg. Podwzgórze jest ograniczone w części przez wyspecjalizowane regiony mózgu, które nie mają skutecznej bariery krew-mózg; śródbłonek kapilar w tych miejscach jest fenestrowany, aby umożliwić swobodne przejście nawet dużych białek i innych cząsteczek. Niektóre z tych miejsc to miejsca neurosekrecji – neurohypophysis i eminencja przyśrodkowa. Jednak inne są miejscami, w których mózg pobiera próbki składu krwi. Dwa z tych miejsc, narząd podoczodołowy i OVLT (organum vasculosum of the lamina terminalis) są tzw. narządami okrężnymi, w których neurony pozostają w ścisłym kontakcie zarówno z krwią, jak i płynem mózgowo-rdzeniowym. Struktury te są gęsto unaczynione i zawierają neurony osmoreceptorowe i sodoreceptorowe, które kontrolują picie, uwalnianie wazopresyny, wydalanie sodu i apetyt na sód. Zawierają one również neurony z receptorami dla angiotensyny, przedsionkowego czynnika natriuretycznego, endoteliny i relaksyny, z których każdy jest ważny w regulacji równowagi płynów i elektrolitów. Neurony w OVLT i SFO rzutują się na jądro nadwzrokowe i jądro przyśrodkowe, a także na obszary podwzgórza przedwzrokowego. Narządy okołokomorowe mogą być również miejscem działania interleukin w celu wywołania zarówno gorączki, jak i wydzielania ACTH, poprzez wpływ na neurony przyśrodkowe.
Nie jest jasne, jak wszystkie peptydy, które wpływają na aktywność podwzgórza, uzyskują niezbędny dostęp. W przypadku prolaktyny i leptyny istnieją dowody na aktywny wychwyt w splocie naczyniówkowym z krwi do płynu mózgowo-rdzeniowego. Niektóre hormony przysadki mózgowej mają ujemne sprzężenie zwrotne na wydzielanie podwzgórzowe; na przykład hormon wzrostu działa na podwzgórze, ale w jaki sposób dostaje się do mózgu, nie jest jasne. Istnieją również dowody na centralne działania prolaktyny i TSH.
Steroidy
Podwzgórze zawiera neurony, które są wrażliwe na steroidy gonadalne i glikokortykoidy – (hormony steroidowe nadnerczy, uwalniane w odpowiedzi na ACTH). Zawiera również wyspecjalizowane neurony wrażliwe na glukozę (w jądrze łukowatym i podwzgórzu brzusznym), które są ważne dla apetytu. Okolica przedwzrokowa zawiera neurony termowrażliwe; są one ważne dla wydzielania TRH.
Wejścia neuronalne
Podwzgórze otrzymuje wiele sygnałów z pnia mózgu; szczególnie z jądra drogi samotnej, jądra współczulnego i rdzenia brzusznego. Wydzielanie oksytocyny w odpowiedzi na ssanie lub stymulację pochwowo-szyjkową odbywa się za pośrednictwem niektórych z tych dróg; wydzielanie wazopresyny w odpowiedzi na bodźce sercowo-naczyniowe pochodzące z chemoreceptorów w zatoce szyjnej i łuku aorty oraz z niskociśnieniowych receptorów objętościowych w przedsionku odbywa się za pośrednictwem innych. U szczura stymulacja pochwy również powoduje wydzielanie prolaktyny, co skutkuje pseudociążą po niepłodnym kryciu. U królika, współżycie wywołuje odruchową owulację. U owcy, stymulacja szyjki macicy w obecności wysokich poziomów estrogenów może wywołać zachowania macierzyńskie u dziewiczej owcy. Wszystkie te efekty pośredniczone są przez podwzgórze, a informacje przenoszone są głównie drogami rdzeniowymi, które znajdują się w pniu mózgu. Stymulacja sutków stymuluje uwalnianie oksytocyny i prolaktyny i tłumi uwalnianie LH i FSH.
Bodźce sercowo-naczyniowe są przenoszone przez nerw błędny, ale błędny przekazuje również różne informacje trzewne, w tym na przykład sygnały wynikające z żołądka rozciągnięcia do tłumienia karmienia. Ponownie te informacje docierają do podwzgórza poprzez przekaźniki w pniu mózgu.
Jądra
Jądra podwzgórza obejmują następujące elementy:
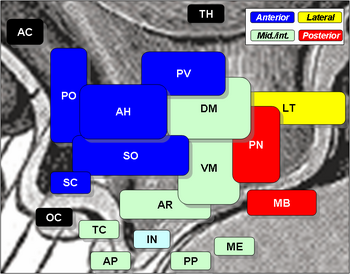
Jądra podwzgórza
| Region | Obszar | Jądro | Jądro | Funkcja |
| Jądro przednie | Jądro przyśrodkowe | Jądro przedoptyczne przyśrodkowe |
|
|
| Jądro przedwzrokowe (SO) jądro nadskrzyżowaniowe (SO) |
|
|||
| jądro przyśrodkowe (PV) |
|
|||
| przednie jądro podwzgórza (AH) |
|
|||
| Jądro nadskrzyżowaniowe (SC) |
|
|||
| Jądro boczne | Jądro przedoptyczne boczne | |||
| Jądro boczne (LT) |
|
|||
| Część jądra nadoptycznego (SO) |
|
|||
| Jądro trzewne | Jądro przyśrodkowe | Jądro grzbietowo-przyśrodkowe podwzgórza (DM) |
|
|
| Jądro przednio-przyśrodkowe (VM) |
. |
|||
Jądro łukowate (AR)
|
||||
| Jądro boczne | . Jądro boczne (LT) |
|
||
| Jądra kanalikowe boczne | ||||
| Jądra tylne | Jądra przyśrodkowe | Jądra migdałowate (część ciałek ssaczych) (MB) |
|
|
| Jądro tylne (PN) |
|
|||
| Jądro boczne (LT) |
Zobacz także: ventrolateral preoptic nucleus
Wyjścia
Wyjścia podwzgórza można podzielić na dwie kategorie: projekcje neuronalne, oraz hormony endokrynne.
Projekcje neuronalne
Większość systemów włókien podwzgórza przebiega w dwóch kierunkach (dwukierunkowo).
- Projekcje do obszarów położonych ogonowo do podwzgórza przechodzą przez pęczek przyśrodkowy przodomózgowia, trakt mammillotegmentalny i powięź podłużną grzbietową.
- Projekcje do obszarów rostralnych do podwzgórza są przenoszone przez przewód mammillothalamic, fornix i terminal stria.
Hormony endokrynne
Podwzgórze wpływa na układ endokrynny i reguluje zachowania emocjonalne, takie jak, gniew i aktywność seksualna. Większość z podwzgórza hormony generowane są dystrybuowane do przysadki mózgowej przez podwzgórza systemu portalowego. Podwzgórze utrzymuje homeostazę, co obejmuje regulację ciśnienia krwi, tętna i temperatury.
Podstawowymi hormonami podwzgórza są:
| Nazwa | Inne nazwy | Skróty | Miejsce | Funkcja |
|---|---|---|---|---|
| Hormon uwalniający kortykotropinę | Faktor uwalniający kortykotropinę, kortykoliberyna | CRH, CRF | przykomórkowe neurony neuroendokrynne w jądrze przyśrodkowym | z wazopresyną, pobudza przednią przysadkę do wydzielania ACTH |
| Dopamina | Hormon hamujący prolaktynę | DA, PIH | neuroendokrynne neurony jądra łukowatego | hamuje wydzielanie prolaktyny z przedniej części przysadki |
| Hormon uwalniający gonadotropiny | Hormon uwalniający hormon luteinizujący | GnRH, LHRH | neuroendokrynne neurony w przyśrodkowym jądrze przedoptycznym i jądrze łukowatym | stymuluje przednią przysadkę do wydzielania LH i FSH |
| Hormon uwalniający hormon wzrostu | Faktor uwalniający hormon wzrostu, somatokrynina | GHRH, GHRF, GRF | neurony neuroendokrynne jądra łukowatego | stymuluje przednią przysadkę do wydzielania hormonu wzrostu |
| Melatonina | Jądro nadskrzyżowaniowe | |||
| Somatostatyna | Hormon wzrostu.hormon hamujący, Czynnik hamujący uwalnianie somatotropiny | SS, GHIH, SRIF | neuroendokrynne neurony jądra okołokomorowego | zahamowanie wydzielania hormonu wzrostu z przedniej części przysadki mózgowej |
| Hormon uwalniający tyreotropinę | Faktor uwalniający tyreotropinę, Thyroliberin, Protirelin | TRH, TRF | parvocellular neuroendocrine neurons in the paraventricular and anterior hypothalamic nuclei | stimulates anterior pituitary to secrete TSH |
See also: Hipokretyna
Kontrola przyjmowania pokarmów
Skrajnie boczna część jądra brzuszno-przyśrodkowego podwzgórza jest odpowiedzialna za kontrolę przyjmowania pokarmów. Stymulacja tego obszaru powoduje zwiększone spożycie pokarmu. Obustronne uszkodzenie tego obszaru powoduje całkowite zaprzestanie przyjmowania pokarmu. Przyśrodkowe części jądra mają kontrolujący wpływ na część boczną. Obustronne uszkodzenie przyśrodkowej części jądra brzuszno-przyśrodkowego powoduje hiperfagię i otyłość zwierzęcia. Dalsze uszkodzenie bocznej części jądra brzusznego u tego samego zwierzęcia powoduje całkowite zaprzestanie przyjmowania pokarmu.
Istnieją różne hipotezy związane z tą regulacją:
- Hipostatyczna hipoteza – ta hipoteza utrzymuje, że tkanka tłuszczowa wytwarza humoralny sygnał, który jest proporcjonalny do ilości tłuszczu i działa na podwzgórze w celu zmniejszenia przyjmowania pokarmu i zwiększenia produkcji energii. Zostało udowodnione, że hormon leptyny działa na podwzgórze w celu zmniejszenia spożycia żywności i zwiększenia produkcji energii.
- Hipoteza jelitowo-peptydowa – hormony przewodu pokarmowego, takie jak Grp, glukagony, CCK i inne twierdził, że hamują spożycie żywności. Jedzenie wchodzące do przewodu pokarmowego wyzwala uwalnianie tych hormonów, które działają na mózg w celu wytworzenia sytości. W mózgu znajdują się zarówno receptory CCK-A, jak i CCK-B.
- Hipoteza glukostatyczna – aktywność ośrodka sytości w jądrach przykomorowych jest prawdopodobnie regulowana przez utylizację glukozy w neuronach. Postuluje się, że gdy ich wykorzystanie glukozy jest niskie i w konsekwencji, gdy różnica glukozy w tętniczo-żylnej krwi przez nie jest niska, aktywność neuronów spada. W tych warunkach aktywność ośrodka żerowania jest niekontrolowana, a osoba odczuwa głód. Przyjmowanie pokarmu jest gwałtownie zwiększane przez dokomorowe podanie 2-deoksyglukozy, co zmniejsza wykorzystanie glukozy w komórkach.
- Hipoteza termostatyczna – według tej hipotezy spadek temperatury ciała poniżej danego punktu nastawczego pobudza apetyt, natomiast wzrost powyżej punktu nastawczego hamuje apetyt.
Dymorfizm płciowy
Kilka jąder podwzgórza jest dymorficznych płciowo, tzn. istnieją wyraźne różnice zarówno w strukturze, jak i funkcji między samcami i samicami.
Niektóre różnice są widoczne nawet w neuroanatomii brutto: najbardziej godne uwagi jest jądro dymorficzne płciowo w obrębie okolicy preoptycznej, które jest obecne tylko u samców. Jednak większość różnic to subtelne zmiany w łączności i wrażliwości chemicznej poszczególnych zestawów neuronów.
Ważność tych zmian można rozpoznać po różnicach funkcjonalnych między samcami i samicami. Na przykład, wzór wydzielania hormonu wzrostu jest dymorficzny płciowo i jest to jeden z powodów, dla których u wielu gatunków dorosłe samce są znacznie większe od samic.
Odpowiedzi na sterydy jajnikowe
Inne uderzające dymorfizmy funkcjonalne dotyczą odpowiedzi behawioralnych na sterydy jajnikowe u dorosłych. Mężczyźni i kobiety różnie reagują na steroidy jajnikowe, częściowo dlatego, że ekspresja wrażliwych na estrogeny neuronów w podwzgórzu jest dymorficzna płciowo, tzn. receptory estrogenowe ulegają ekspresji w różnych zestawach neuronów.
Estrogeny i progesteron mogą wpływać na ekspresję genów w poszczególnych neuronach lub wywoływać zmiany w potencjale błony komórkowej i aktywacji kinaz, prowadząc do różnorodnych niegenomowych funkcji komórkowych. Estrogen i progesteron wiążą się ze swoimi receptorami hormonów jądrowych, które przemieszczają się do jądra komórkowego i oddziałują z regionami DNA znanymi jako elementy odpowiedzi hormonalnej (HRE) lub wiążą się z miejscem wiązania innego czynnika transkrypcyjnego. Wykazano, że receptor estrogenowy (ER) może w ten sposób tranaktywować inne czynniki transkrypcyjne, pomimo braku elementu odpowiedzi estrogenowej (ERE) w proksymalnym regionie promotorowym genu. ERs i receptory progesteronu (PRs) są generalnie aktywatorami genów, ze zwiększonym mRNA i późniejszą syntezą białka po ekspozycji hormonalnej.
Mózgi mężczyzn i kobiet różnią się w rozmieszczeniu receptorów estrogenowych, a różnica ta jest nieodwracalną konsekwencją noworodkowej ekspozycji na sterydy. Receptory estrogenowe (i progesteronowe) znajdują się głównie w neuronach w przednim i środkowym podwzgórzu, w szczególności:
- obszarze preoptycznym (gdzie znajdują się neurony LHRH)
- jądrze okołokomorowym (gdzie znajdują się neurony somatostatynowe)
- przyśrodkowym podwzgórzu (które jest ważne dla zachowań seksualnych).
Steroidy gonadalne w życiu noworodkowym szczurów
W życiu noworodkowym steroidy gonadalne wpływają na rozwój neuroendokrynny podwzgórza. Na przykład, określają one zdolność samic do wykazywania normalnego cyklu reprodukcyjnego oraz samców i samic do wykazywania odpowiednich zachowań reprodukcyjnych w życiu dorosłym.
- Jeśli samica szczura jest wstrzykiwany raz z testosteronem w ciągu pierwszych kilku dni życia postnatalnego (podczas „okresu krytycznego” wpływu sex-steroidów), podwzgórze jest nieodwracalnie maskulinized; dorosły szczur będzie niezdolny do generowania skoku LH w odpowiedzi na estrogen (charakterystyczne dla samic), ale będzie w stanie wykazać męskich zachowań seksualnych (montaż płciowo receptywnej samicy).
- Przez kontrast, samiec szczura wykastrowany tuż po urodzeniu będzie feminizowany, a dorosły będzie wykazywał żeńskie zachowania seksualne w odpowiedzi na estrogen (receptywność seksualna, lordoza).
Androgeny w naczelnych
W naczelnych, rozwojowy wpływ androgenów jest mniej jasne, a konsekwencje są mniej kompletne. Tomboyism” w dziewcząt może odzwierciedlać skutki androgenów na płodowym mózgu, ale płeć wychowania w ciągu pierwszych 2-3 lat jest uważany przez wielu, aby być najważniejszym czynnikiem determinującym tożsamości płciowej, ponieważ w tej fazie albo estrogenów lub testosteronu będą miały trwałe skutki albo żeński lub męski mózg, wpływając zarówno heteroseksualności i homoseksualności.
Paradoksem jest to, że maskulinizujące skutki testosteronu są pośredniczone przez estrogen. W mózgu, testosteron jest aromatyzowane do (estradiol), który jest głównym hormonem aktywnym dla wpływów rozwojowych. Ludzkie jądro wydziela wysoki poziom testosteronu od około 8 tygodnia życia płodowego do 5-6 miesiąca po urodzeniu (podobny okołoporodowy wzrost testosteronu obserwuje się u wielu gatunków), proces, który wydaje się leżeć u podstaw męskiego fenotypu. Estrogen z krążenia matki jest stosunkowo nieskuteczne, częściowo z powodu wysokich krążących poziomów białek wiążących steroidy w pregnancy.
Inne wpływy na rozwój podwzgórza
Sex steroidy nie są jedynymi ważnymi wpływami na rozwój podwzgórza, w szczególności, pre-pubertal stres we wczesnym życiu określa zdolność dorosłego podwzgórza do odpowiedzi na ostry stresor. W przeciwieństwie do gonadalnych receptorów steroidowych, receptory glikokortykoidowe są bardzo rozpowszechnione w całym mózgu; w jądrze przyśrodkowym pośredniczą one w kontroli syntezy i wydzielania CRF za pomocą ujemnego sprzężenia zwrotnego, ale w innych miejscach ich rola nie jest dobrze poznana.
Wpływ starzenia się na podwzgórze
Badania na samicach myszy wykazały, że zarówno jądro nadoptyczne (SON), jak i jądro przyśrodkowe (PVN) tracą około jednej trzeciej immunoreaktywnych komórek IGF-1R przy normalnym starzeniu się. Ponadto, myszy Old-CR utraciły większą liczbę nieimmunoreaktywnych komórek IGF-1R przy zachowaniu podobnej liczby komórek immunoreaktywnych IGF-1R w porównaniu do myszy Old-Al. W konsekwencji, myszy Old-CR wykazują wyższy odsetek komórek immunoreaktywnych IGF-1R, co odzwierciedla zwiększoną wrażliwość podwzgórza na IGF-1 w porównaniu do normalnie starzejących się myszy.
Zobacz także
- Oś HPA
- Układ podwzgórzowo-przysadkowo-nadnerczowy
- Uszkodzenia podwzgórza
- Pęczek przedni przyśrodkowy
- Neuroendokrynologia
- Obszar przedwzrokowy
.
Obrazy dodatkowe
Dodaj zdjęcie do tej galerii
- http://www.cancer.gov/Templates/db_alpha.aspx?CdrID=46359
- http://www.sci.uidaho.edu/med532/hypothal.htm
- Diagram jąder (psycheducation.org)
- Diagram of Nuclei (universe-review.ca)
- Diagram of Nuclei (utdallas.edu)
- Unless else specified in table, then ref is: Guyton Eight Edition
- Walter F., PhD. Boron. Medical Physiology: A Cellular And Molecular Approaoch, Elsevier/Saunders. Strona 840
- http://thalamus.wustl.edu/course/hypoANS.html
- http://www.vivo.colostate.edu/hbooks/pathphys/endocrine/hypopit/overview.html
- Theologides A (1976). Anorexia-producing metabolitów pośrednich. Am J Clin Nutr 29 (5): 552-8.
- John Money, „The concept of gender identity disorder in childhood and adolescence after 39 years”, Journal of Sex and Marital Therapy 20 (1994): 163-77.
- Romeo, Russell D, Rudy Bellani, Ilia N. Karatsoreos, Nara Chhua, Mary Vernov, Cheryl D. Conrad and Bruce S. McEwen (2005). Historia stresu i Pubertal rozwoju interakcji Shape Plasticity Hypothalamic-Pituitary-Adrenal Axis. Endocrinology 147 (4): 1664-1674.
- Saeed O,Yaghmaie F,Garan SA,Gouw AM,Voelker MA,Sternberg H, Timiras PS. (2007). Insulin-like growth factor-1 receptor immunoreactive cells are selectively maintained in the paraventricular hypothalamus of calorically restricted mice. Int J Dev Neurosci 25 (1): 23-8.
- Yaghmaie F, Saeed O, Garan SA, Voelker MA, Gouw AM, Freitag W, Sternberg H, Timiras PS (2006). Age-dependent loss of insulin-like growth factor-1 receptor immunoreactive cells in the supraoptic hypothalamus is reduced in calorically restricted mice. Int J Dev Neurosci 24 (7): 431-6.
- F. Yaghmaie, O. Saeed, S.A. Garan, A.M. Gouw, P. Jafar, J. Kaur, S. Nijjar, P.S. Timiras, H. Sternberg, M.A. Voelker (2007). Śledzenie zmian w podwzgórzu IGF-1 wrażliwości z starzenia się i kalorycznej restrykcji. Experimental Gerontology 42 (1-2): 148-149.
Referencje & Bibliografia
Prace
- Arees, E.A. i Mayer, B, (1967) Anatomiczne połączenia między przyśrodkowymi i bocznymi regionami podwzgórza związane z przyjmowaniem pokarmu, Science 157: 1574-5
- Grossman, S.P. (1960) Eating or drinking elicited by direct adrenergic or cholinergic stimulation of the hypothalamus, Science 132: 301-2.
- Hetherington, A.W. and Ranson, S.W. (1942) The relation of various hypothalamic lesions to adiposity in the rat, 76: 475-99.
Materiały dodatkowe
Papiery
- Google Scholar
.
- BrainMaps at UCDavis Hypothalamus
- Układ dokrewny i podwzgórze
- High-Resolution Cytoarchitectural Primate Brain Atlases
- The Hypothalamus and Pituitary at endotexts.org
- Diagram of Nuclei (psycheducation.org)
- Diagram of Nuclei (universe-review.ca)
- Diagram of Nuclei (utdallas.edu)
|
v-d-e
Mózg człowieka: diencephalon (TA A14.1.08, GA 9.807) |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Epithalamus |
|
Powierzchnia
|
||||||||||||||
| Wzgórze |
|
Powierzchnia
|
Substancja szara/
jądra |
|||||||||||||
| Hypothalamus |
|
|||||||||||||||
| Podwzgórze |
Jądro podwzgórzowe – Zona incerta |
|||||||||||||||
| Układ komorowy: Trzecia komora |
wnęki: (Optic recess, Infundibular recess, Suprapineal recess, Pineal recess) Hypothalamic sulcus – Tela chorioidea of third ventricle Subfornical organ Apertures: Interventricular/Monro Posterior commissure |
|||||||||||||||
|
v-d-e
Anatomia człowieka, układ dokrewny: gruczoły dokrewne |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Osie podwzgórze/ przysadka |
|
|
|
|
||||||||||||
| Gruczoł szyszynki |
Gruczoł szyszynki – Corpora arenacea |
|||||||||||||||
| Wyrostki trzustki |
Komórka alfa – Komórka beta – Komórka delta – Komórka PP – Komórka epsilon |
|||||||||||||||
Mózg człowieka: Układ limbiczny
Amygdala – Cingulate gyrus – Fornicate gyrus – Hipokamp – Hypothalamus – Mammillary body – Nucleus accumbens – Orbitofrontal cortex – Parahippocampal gyrus
.