Assessment | Biopsychology | Comparative |Cognitive | Developmental | Language | Individual differences |Personality | Philosophy | Social |
Methods | Statistics |Clinical | Educational | Industrial |Professional items |World psychology |
Biological:Gedragsgenetica – Evolutionaire psychologie – Neuroanatomie – Neurochemie – Neuroendocrinologie -Neurowetenschappen – Psychoneuroimmunologie – Fysiologische psychologie – Psychofarmacologie(Index, Outline)
| Hersenen: Hypothalamus | ||
|---|---|---|
 |
||
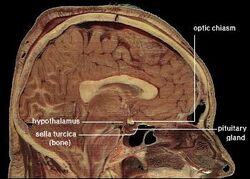
| Locatie van de menselijke hypothalamus | ||
 |
||
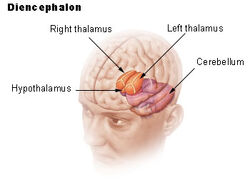
| Dienchephalon | ||
| Latijn | hypothalamus | |
| Gray’s | onderwerp #189 812 | |
| Deel van | ||
| Componenten | ||
| Arterie | ||
| Vein | ||
| BrainInfo/UW | hier-358 | |
| MeSH | A08.186.211.730.385.357 | |
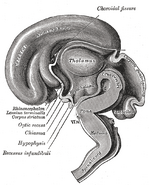
De hypothalamus verbindt het zenuwstelsel met het endocriene stelsel via de hypofyse (hypofyse). De hypothalamus, (van het Griekse ὑποθαλαμος = onder de thalamus) bevindt zich onder de thalamus, net boven de hersenstam. Deze klier beslaat het grootste deel van het ventrale gebied van het diencephalon. Hij komt voor in de hersenen van alle zoogdieren. Bij de mens is zij ongeveer zo groot als een amandel.
De hypothalamus regelt bepaalde stofwisselingsprocessen en andere activiteiten van het Autonoom Zenuwstelsel. Hij synthetiseert en scheidt neurohormonen af, die vaak hypothalamus-releasinghormonen worden genoemd, en deze stimuleren of remmen op hun beurt de secretie van hypofysehormonen.
De hypothalamus regelt de lichaamstemperatuur, honger, dorst, vermoeidheid, woede, en de circadiane cycli.
- Inputs
- Olfactorische prikkels
- Prikkels door het bloed
- Steroïden
- Neurale inputs
- Nuclei
- Uitgangen
- Neurale projecties
- Endocriene hormonen
- Regeling van voedselinname
- Seksueel dimorfisme
- Reacties op ovariële steroïden
- Gonadale steroïden in het neonatale leven van ratten
- Androgenen bij primaten
- Andere invloeden op de ontwikkeling van de hypothalamus
- Effecten van veroudering op de hypothalamus
- Zie ook
- Aanvullende beelden
- Referenties & Bibliografie
- Papers
- Aanvullend materiaal
- Papers
Inputs
De hypothalamus is een zeer complex gebied in de hersenen van de mens, en zelfs kleine kernen binnen de hypothalamus zijn betrokken bij veel verschillende functies. De paraventriculaire kern bijvoorbeeld bevat oxytocine en vasopressine (ook wel antidiuretisch hormoon genoemd) neuronen die naar de achterste hypofyse projecteren, maar bevat ook neuronen die ACTH en TSH secretie reguleren (die naar de voorste hypofyse projecteren), maagreflexen, moederlijk gedrag, bloeddruk, voeding, immuunreacties, en temperatuur.
De hypothalamus coördineert vele hormonale en gedragsmatige circadiane ritmen, complexe patronen van neuro-endocriene outputs, complexe homeostatische mechanismen, en vele belangrijke gedragingen. De hypothalamus moet daarom reageren op veel verschillende signalen, waarvan sommige van buitenaf en andere van binnenuit worden opgewekt. De hypothalamus is dus nauw verbonden met vele delen van het CZS, met inbegrip van de hersenstam reticulaire formatie en autonome zones, de limbische voorhersenen (in het bijzonder de amygdala, het septum, de diagonale band van Broca en de olfactorische bollen, en de hersenschors).
De hypothalamus is gevoelig voor:
- Licht: daglengte en fotoperiode voor het reguleren van circadiane en seizoensgebonden ritmen
- Olfactorische stimuli, waaronder feromonen
- Steroïden, waaronder gonadale steroïden en corticosteroïden
- Natuurlijk doorgegeven informatie die met name afkomstig is van het hart, de maag, en het voortplantingskanaal
- Autonome inputs
- Bloedgestuurde stimuli, waaronder leptine, ghreline, angiotensine, insuline, hypofysaire hormonen, cytokinen, plasmaconcentraties van glucose en osmolariteit enz
- Stress
- Binnenstromende micro-organismen door verhoging van de lichaamstemperatuur, waardoor de lichaamsthermostaat naar boven wordt teruggesteld.
Olfactorische prikkels
Olfactorische prikkels zijn belangrijk voor de voortplanting en de neuro-endocriene functie bij veel diersoorten. Als bijvoorbeeld een zwangere muis wordt blootgesteld aan de urine van een ‘vreemd’ mannetje tijdens een kritieke periode na de coïtus dan mislukt de zwangerschap (het Bruce-effect). Zo vormt een vrouwtjesmuis tijdens de coïtus een nauwkeurig “olfactorisch geheugen” van haar partner, dat verscheidene dagen blijft bestaan.Feromonale signalen helpen bij de synchronisatie van de bronst bij vele soorten; bij vrouwen kan de gesynchroniseerde menstruatie ook het gevolg zijn van feromonale signalen, hoewel de rol van feromonen bij de mens door sommigen wordt betwijfeld.
Prikkels door het bloed
Peptide hormonen hebben belangrijke invloeden op de hypothalamus, en om dit te doen moeten zij de bloed-hersenbarrière omzeilen. De hypothalamus wordt gedeeltelijk begrensd door gespecialiseerde hersengebieden waar geen effectieve bloed-hersenbarrière bestaat; het capillaire endotheel op deze plaatsen is gefenestreerd om zelfs grote proteïnen en andere moleculen vrije doorgang te verlenen. Sommige van deze plaatsen zijn de plaatsen van neurosecretie – de neurohypofyse en de mediane eminentie. Andere zijn echter plaatsen waar de hersenen de samenstelling van het bloed bemonsteren. Twee van deze plaatsen, het subfornicale orgaan en het OVLT (organum vasculosum van de lamina terminalis) zijn zogenaamde circumventriculaire organen, waar de neuronen in nauw contact staan met zowel het bloed als de liquor. Deze structuren zijn sterk gevasculariseerd en bevatten osmoreceptieve en natriumreceptieve neuronen die het drinken, de vasopressine-afgifte, de natriumuitscheiding en de natriumhonger regelen. Zij bevatten ook neuronen met receptoren voor angiotensine, atriale natriuretische factor, endotheline en relaxine, die elk belangrijk zijn voor de regulering van de vocht- en elektrolytenbalans. Neuronen in de OVLT en SFO projecteren naar de supraoptische kern en de paraventriculaire kern, en ook naar preoptische hypothalamische gebieden. De circumventriculaire organen kunnen ook de plaats zijn waar interleukines werken om zowel koorts als ACTH secretie uit te lokken, via effecten op paraventriculaire neuronen.
Het is niet duidelijk hoe alle peptiden die de hypothalamische activiteit beïnvloeden de nodige toegang krijgen. In het geval van prolactine en leptine zijn er aanwijzingen voor actieve opname bij de plexus choroideus vanuit het bloed in de CSF. Sommige hypofysehormonen hebben een negatieve feedback-invloed op de hypothalamische secretie; groeihormoon bijvoorbeeld werkt terug op de hypothalamus, maar hoe het de hersenen binnenkomt is niet duidelijk. Er zijn ook aanwijzingen voor centrale acties van prolactine en TSH.
Steroïden
De hypothalamus bevat neuronen die gevoelig zijn voor gonadale steroïden en glucocorticoïden – (de steroïdhormonen van de bijnier, die vrijkomen als reactie op ACTH). Het bevat ook gespecialiseerde glucosegevoelige neuronen (in de arcuate kern en de ventromediale hypothalamus), die belangrijk zijn voor de eetlust. Het preoptische gebied bevat thermogevoelige neuronen; deze zijn belangrijk voor de TRH secretie.
Neurale inputs
De hypothalamus ontvangt veel inputs van de hersenstam; met name van de nucleus van het solitaire tractus, de locus coeruleus, en de ventrolaterale medulla. De secretie van oxytocine als reactie op zuigen of vaginocervicale stimulatie wordt door sommige van deze wegen bemiddeld; de secretie van vasopressine als reactie op cardiovasculaire stimuli die afkomstig zijn van chemoreceptoren in de sinus carotis en de aortaboog, en van atriale volume-receptoren onder lage druk, wordt door andere bemiddeld. Bij de rat veroorzaakt stimulatie van de vagina ook prolactine secretie, en dit resulteert in pseudo-zwangerschap na een onvruchtbare paring. Bij het konijn veroorzaakt coïtus een reflexmatige ovulatie. Bij het schaap kan stimulatie van de baarmoederhals in aanwezigheid van hoge oestrogeenniveaus moederlijk gedrag bij een maagdelijk ooi opwekken. Al deze effecten worden bemiddeld door de hypothalamus, en de informatie wordt hoofdzakelijk doorgegeven via spinale paden die zich in de hersenstam bevinden. Stimulatie van de tepels stimuleert de afgifte van oxytocine en prolactine en onderdrukt de afgifte van LH en FSH.
Cardiovasculaire prikkels worden overgebracht door de nervus vagus, maar de nervus vagus geeft ook een verscheidenheid aan viscerale informatie door, waaronder bijvoorbeeld signalen die het gevolg zijn van een gespannen maag om het voeden te onderdrukken. Ook deze informatie bereikt de hypothalamus via relais in de hersenstam.
Nuclei
De hypothalamische kernen omvatten de volgende:
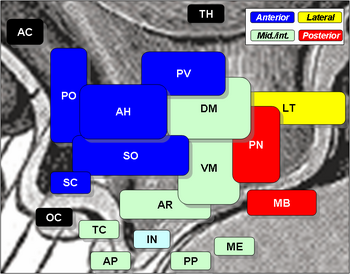
Hypothalamische kernen
| Regio | Area | Nucleus | Functie |
| Anterior | Medial | Medial preoptic nucleus |
|
| Supraoptic nucleus (SO) |
|
||
| Paraventriculaire nucleus (PV) |
|
||
| Anterior hypothalamic nucleus (AH) |
|
||
| Suprachiasmatische kern (SC) |
|
||
| Laterale | Laterale preoptische nucleus | ||
| Laterale nucleus (LT) |
|
||
| Deel van supraoptische nucleus (SO) |
|
||
| Tuberale | Mediale | Dorsomediale hypothalamuskern (DM) |
|
| Ventromediale nucleus (VM) |
|
||
Arcuate nucleus (AR)
|
|||
| Laterale | Laterale nucleus (LT) |
|
|
| Laterale tuberale kernen | |||
| Posterior | Medial | Mammillaire kernen (deel van mammillaire lichaampjes) (MB) |
|
| Posterior nucleus (PN) |
|
||
| Laterale | Laterale nucleus (LT) |
Zie ook: ventrolaterale preoptische kern
Uitgangen
De uitgangen van de hypothalamus kunnen in twee categorieën worden onderverdeeld: neurale projecties, en endocriene hormonen.
Neurale projecties
De meeste vezelstelsels van de hypothalamus lopen in twee richtingen (bidirectioneel).
- Projecties naar gebieden caudaal van de hypothalamus gaan via de mediale voorhersenenbundel, de mammillotegmentale tractus en de dorsale longitudinale fasciculus.
- Projecties naar gebieden rostraal van de hypothalamus gaan door de mammillothalamus tractus, de fornix en de terminale stria.
Endocriene hormonen
De hypothalamus beïnvloedt het endocriene systeem en regelt emotioneel gedrag, zoals, woede en seksuele activiteit. De meeste hormonen die door de hypothalamus worden geproduceerd, worden via het hypophyseale portaalsysteem naar de hypofyse gedistribueerd. De hypothalamus handhaaft de homeostase, hetgeen de regulering van de bloeddruk, de hartslag en de temperatuur omvat.
De primaire hypothalamushormonen zijn:
| Naam | Andere namen | Afkortingen | Locatie | Functie |
|---|---|---|---|---|
| Corticotropin-releasing hormone | Corticotropin-releasing factor, Corticoliberin | CRH, CRF | parvocellulaire neuroendocriene neuronen in de paraventriculaire kern | met vasopressine, stimuleert anterieure hypofyse tot afscheiding van ACTH |
| Dopamine | Prolactine-remmend hormoon | DA, PIH | neuroendocriene neuronen van de arcuate kern | remt secretie van prolactine uit de voorste hypofyse |
| Gonadotropin-releasing hormone | Luteinising-hormone releasing hormone | GnRH, LHRH | neuro-endocriene neuronen in de mediale preoptische en arcuate kernen | stimuleert de voorste hypofyse tot de afscheiding van LH en FSH |
| Groeihormoon-afgiftehormoon | Groeihormoon-afgiftefactor, somatocrinine | GHRH, GHRF, GRF | arcuate nucleus neuroendocriene neuronen | stimuleert anterieure hypofyse tot afscheiding van groeihormoon |
| Melatonine | suprachiasmatische kern | |||
| Somatostatine | Groeihormoon-remmend hormoon, Somatotropine-afgifte-remmende factor | SS, GHIH, SRIF | neuro-endocriene neuronen van de periventriculaire kern | remt secretie van groeihormoon uit de voorste hypofyse |
| Thyrotropine-afgiftehormoon | Thyrotropine-afgiftefactor, Thyroliberine, Protireline | TRH, TRF | parvocellulaire neuroendocriene neuronen in de paraventriculaire en anterieure hypothalamische kernen | stimuleert anterieure hypofyse om TSH af te scheiden |
Zie ook: Hypocretine
Regeling van voedselinname
Het uiterste laterale deel van de ventromediale kern van de hypothalamus is verantwoordelijk voor de regeling van de voedselinname. Stimulatie van dit gebied veroorzaakt een verhoogde voedselinname. Bilaterale laesie van dit gebied veroorzaakt volledige stopzetting van de voedselinname. Mediale delen van de kern hebben een controlerend effect op het laterale deel. Bilaterale lesie van het mediale deel van de ventromediale nucleus veroorzaakt hyperfagie en zwaarlijvigheid bij het dier. Verdere laesie van het laterale deel van de ventromediale kern in hetzelfde dier veroorzaakt volledige stopzetting van de voedselinname.
Er zijn verschillende hypothesen met betrekking tot deze regulatie:
- Lipostatische hypothese – deze hypothese stelt dat vetweefsel een humoraal signaal produceert dat evenredig is met de hoeveelheid vet en inwerkt op de hypothalamus om de voedselinname te verminderen en de energie-output te verhogen. Het is gebleken dat het hormoon leptine op de hypothalamus inwerkt om de voedselinname te verminderen en de energie-output te verhogen.
- Gutpeptide-hypothese – gastro-intestinale hormonen zoals Grp, glucagonen, CCK en andere zouden de voedselinname afremmen. Het voedsel dat het maagdarmkanaal binnenkomt, veroorzaakt de afgifte van deze hormonen die inwerken op de hersenen om verzadiging te produceren. De hersenen bevatten zowel CCK-A als CCK-B receptoren.
- Glucostatische hypothese – de activiteit van het verzadigingscentrum in de ventromediale kernen wordt waarschijnlijk bepaald door de glucosebenutting in de neuronen. Er is verondersteld dat wanneer hun glucosegebruik laag is en bijgevolg wanneer het arterioveneuze bloedglucoseverschil over hen laag is, de activiteit over de neuronen afneemt. Onder deze omstandigheden is de activiteit van het voedingscentrum ongecontroleerd en voelt het individu zich hongerig. De voedselopname wordt snel verhoogd door intraventriculaire toediening van 2-deoxyglucose, waardoor het glucosegebruik in de cellen daalt.
- Thermostatische hypothese – volgens deze hypothese stimuleert een daling van de lichaamstemperatuur onder een bepaald setpoint de eetlust, terwijl een stijging boven het setpoint de eetlust remt.
Seksueel dimorfisme
Verschillende hypothalamische kernen zijn seksueel dimorf, d.w.z. er zijn duidelijke verschillen in zowel structuur als functie tussen mannetjes en vrouwtjes.
Sommige verschillen zijn zelfs in de grove neuro-anatomie duidelijk: het meest opmerkelijk is de seksueel dimorfe kern in het preopticumgebied, die alleen bij mannetjes aanwezig is. De meeste verschillen zijn echter subtiele veranderingen in de connectiviteit en de chemische gevoeligheid van bepaalde groepen neuronen.
Het belang van deze veranderingen kan worden herkend aan de hand van functionele verschillen tussen mannetjes en vrouwtjes. Zo is het patroon van secretie van groeihormoon seksueel dimorf, en dit is een van de redenen waarom bij veel soorten volwassen mannetjes veel groter zijn dan vrouwtjes.
Reacties op ovariële steroïden
Andere opvallende functionele dimorfismen zijn in de gedragsreacties op ovariële steroïden van de volwassene. Mannetjes en vrouwtjes reageren verschillend op ovariumsteroïden, deels omdat de expressie van oestrogeen-gevoelige neuronen in de hypothalamus sexueel dimorf is, d.w.z. oestrogeen-receptoren komen tot expressie in verschillende sets neuronen.
Oestrogeen en progesteron kunnen genexpressie in bepaalde neuronen beïnvloeden of veranderingen induceren in celmembraanpotentiaal en kinase-activatie, wat leidt tot diverse niet-genomische cellulaire functies. Oestrogeen en progesteron binden zich aan hun eigen nucleaire hormoonreceptoren, die zich naar de celkern verplaatsen en daar in wisselwerking staan met DNA-gebieden die bekend staan als hormoonresponselementen (HRE’s) of die worden gebonden aan de bindingsplaats van een andere transcriptiefactor. Van de oestrogeenreceptor (ER) is aangetoond dat hij op deze wijze andere transcriptiefactoren transactiveert, ondanks de afwezigheid van een oestrogeenresponselement (ERE) in de proximale promotorregio van het gen. ER’s en progesteronreceptoren (PR’s) zijn in het algemeen genactivatoren, met een verhoogde mRNA en daaropvolgende eiwitsynthese na blootstelling aan hormonen.
De hersenen van mannen en vrouwen verschillen in de verdeling van oestrogeenreceptoren, en dit verschil is een onomkeerbaar gevolg van neonatale blootstelling aan steroïden. Oestrogeenreceptoren (en progesteronreceptoren) worden voornamelijk aangetroffen in neuronen in de voorste en mediobasale hypothalamus, met name:
- de preoptische zone (waar zich LHRH-neuronen bevinden)
- de periventriculaire kern (waar zich somatostatine-neuronen bevinden)
- de ventromediale hypothalamus (die belangrijk is voor seksueel gedrag).
Gonadale steroïden in het neonatale leven van ratten
In het neonatale leven beïnvloeden gonadale steroïden de ontwikkeling van de neuro-endocriene hypothalamus. Zij bepalen bijvoorbeeld het vermogen van wijfjes om een normale voortplantingscyclus te vertonen, en van mannetjes en wijfjes om in het volwassen leven passend voortplantingsgedrag te vertonen.
- Als een vrouwtjesrat in de eerste dagen van het postnatale leven (tijdens de “kritieke periode” van beïnvloeding door sex-steroïden) eenmaal met testosteron wordt geïnjecteerd, wordt de hypothalamus onomkeerbaar vermannelijkt; de volwassen rat zal niet in staat zijn een LH-piek te genereren als reactie op oestrogeen (een kenmerk van vrouwtjes), maar zal wel in staat zijn mannelijk seksueel gedrag te vertonen (het bestijgen van een seksueel ontvankelijk vrouwtje).
- Daarentegen zal een mannelijke rat die vlak na de geboorte gecastreerd is, vervrouwelijkt zijn, en de volwassene zal vrouwelijk seksueel gedrag vertonen als reactie op oestrogeen (seksuele ontvankelijkheid, lordose).
Androgenen bij primaten
Bij primaten is de invloed van androgenen op de ontwikkeling minder duidelijk, en de gevolgen zijn minder volledig. ‘Tomboyisme’ bij meisjes zou een weerspiegeling kunnen zijn van de effecten van androgenen op het foetale brein, maar het geslacht van de opvoeding gedurende de eerste 2-3 jaar wordt door velen beschouwd als de belangrijkste determinant van geslachtsidentiteit, omdat gedurende deze fase ofwel oestrogeen ofwel testosteron blijvende effecten zal hebben op ofwel een vrouwelijk ofwel een mannelijk brein, en zowel heteroseksualiteit als homoseksualiteit zal beïnvloeden.
De paradox is dat de masculiniserende effecten van testosteron worden gemedieerd door oestrogeen. In de hersenen wordt testosteron gearomatiseerd tot (oestradiol), dat het belangrijkste actieve hormoon is voor de ontwikkelingsbeïnvloeding. De menselijke testis scheidt hoge niveaus testosteron af vanaf ongeveer week 8 van het foetale leven tot 5-6 maanden na de geboorte (een soortgelijke perinatale stijging van testosteron wordt waargenomen bij vele diersoorten), een proces dat ten grondslag lijkt te liggen aan het mannelijk fenotype. Oestrogeen uit de maternale circulatie is relatief ineffectief, deels vanwege de hoge circulerende niveaus van steroïde-bindende proteïnen tijdens de zwangerschap.
Andere invloeden op de ontwikkeling van de hypothalamus
Seksuele steroïden zijn niet de enige belangrijke invloeden op de ontwikkeling van de hypothalamus; met name pre-puberale stress in het vroege leven bepaalt de capaciteit van de volwassen hypothalamus om te reageren op een acute stressor. In tegenstelling tot gonadale steroïdreceptoren zijn glucocorticoïdreceptoren zeer wijdverspreid in de hersenen; in de paraventriculaire kern bemiddelen zij bij de negatieve terugkoppelingscontrole van CRF-synthese en -secretie, maar elders is hun rol niet goed begrepen.
Effecten van veroudering op de hypothalamus
Studies bij vrouwelijke muizen hebben aangetoond dat zowel de Supraoptic nucleus (SON) als de Paraventriculaire nucleus (PVN) ongeveer een derde van de IGF-1R immunoreactieve cellen verliezen bij normale veroudering. Ook verloren oude muizen met calorierestrictie (CR) grotere aantallen niet-immunoreactieve IGF-1R cellen, terwijl ze vergelijkbare aantallen IGF-1R immunoreactieve cellen behielden in vergelijking met oude-Al muizen. Bijgevolg vertonen Oud-CR muizen een hoger percentage IGF-1R immunoreactieve cellen, wat wijst op een verhoogde gevoeligheid van de hypothalamus voor IGF-1 in vergelijking met normaal ouder wordende muizen.
Zie ook
- HPA-as
- Hypothalamo hypofyse systeem
- Hypothalamo hypofyse bijnier systeem
- Hypothalamus laesies
- Mediale voorhersenen bundel
- Neuroendocrinologie
- Preoptisch gebied
Aanvullende beelden
Een foto aan deze galerij toevoegen
- http://www.cancer.gov/Templates/db_alpha.aspx?CdrID=46359
- http://www.sci.uidaho.edu/med532/hypothal.htm
- Schema van kernen (psycheducation.org)
- Diagram of Nuclei (universe-review.ca)
- Diagram of Nuclei (utdallas.edu)
- Tenzij anders vermeld in tabel, dan is ref: Guyton Eight Edition
- Walter F., PhD. Boron. Medische Fysiologie: A Cellular And Molecular Approaoch, Elsevier/Saunders. Pagina 840
- http://thalamus.wustl.edu/course/hypoANS.html
- http://www.vivo.colostate.edu/hbooks/pathphys/endocrine/hypopit/overview.html
- Theologides A (1976). Anorexia-producerende intermediaire metabolieten. Am J Clin Nutr 29 (5): 552-8.
- John Money, ‘The concept of gender identity disorder in childhood and adolescence after 39 years’, Journal of Sex and Marital Therapy 20 (1994): 163-77.
- Romeo, Russell D, Rudy Bellani, Ilia N. Karatsoreos, Nara Chhua, Mary Vernov, Cheryl D. Conrad and Bruce S. McEwen (2005). Stressgeschiedenis en puberteitsontwikkeling beïnvloeden samen de plasticiteit van de hypothalamus-hypofyse-bijnieras. Endocrinology 147 (4): 1664-1674.
- Saeed O,Yaghmaie F,Garan SA,Gouw AM,Voelker MA,Sternberg H, Timiras PS. (2007). Insulin-like growth factor-1 receptor immunoreactive cells are selectively maintained in the paraventricular hypothalamus of calorically restricted mice. Int J Dev Neurosci 25 (1): 23-8.
- Yaghmaie F, Saeed O, Garan SA, Voelker MA, Gouw AM, Freitag W, Sternberg H, Timiras PS (2006). Age-dependent loss of insulin-like growth factor-1 receptor immunoreactive cells in the supraoptic hypothalamus is reduced in calorically restricted mice. Int J Dev Neurosci 24 (7): 431-6.
- F. Yaghmaie, O. Saeed, S.A. Garan, A.M. Gouw, P. Jafar, J. Kaur, S. Nijjar, P.S. Timiras, H. Sternberg, M.A. Voelker (2007). Tracking changes in hypothalamic IGF-1 sensitivity with aging and caloric restriction. Experimentele gerontologie 42 (1-2): 148-149.
Referenties & Bibliografie
Papers
- Arees, E.A. and Mayer, B, (1967) Anatomical connections between medial and lateral regions of the hypothalamus concerned with food intake, Science 157: 1574-5
- Grossman, S.P. (1960) Eating or drinking elicited by direct adrenergic or cholinergic stimulation of the hypothalamus, Science 132: 301-2.
- Hetherington, A.W. and Ranson, S.W. (1942) The relation of various hypothalamic lesions to adiposity in the rat, 76: 475-99.
Aanvullend materiaal
Papers
- Google Scholar
- BrainMaps at UCDavis Hypothalamus
- Endocrien systeem en hypothalamus
- High-Resolution Cytoarchitectural Primate Brain Atlases
- The Hypothalamus and Pituitary at endotexts.org
- Diagram van Nuclei (psycheducation.org)
- Diagram van Nuclei (universe-review.ca)
- Diagram van Nuclei (utdallas.edu)
|
v-d-e
Menselijk brein: diencephalon (TA A14.1.08, GA 9.807) |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Epithalamus |
|
Surface
|
||||||||||||||
| Thalamus |
|
Surface
|
Grijze stof/
nuclei |
|||||||||||||
| Hypothalamus |
|
|||||||||||||||
| Subthalamus |
Subthalamuskern – Zona incerta |
|||||||||||||||
| Ventriculair systeem: Derde ventrikel |
uitsparingen: (Optische uitsparing, Infundibulaire uitsparing, Suprapineale uitsparing, Pijnappeluitsparing) Hypothalamische sulcus – Tela chorioidea van derde ventrikel Subfornicale orgaan Aperturen: Interventriculair/Monro Posterior commissure |
|||||||||||||||
|
v-d-e
Menselijke anatomie, endocrien systeem: endocriene klieren |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hypothalamus/ hypofyse-assen |
|
|
|
|
||||||||||||
| Pineale klier |
Pinealocyten – Corpora arenacea |
|||||||||||||||
| Islets van pancreas |
Alpha cel – Beta cel – Delta cel – PP cel – Epsilon cel |
|||||||||||||||
Hersenen van de mens: Limbisch systeem
Amygdala – Cingulate gyrus – Fornicate gyrus – Hippocampus – Hypothalamus – Mammillary body – Nucleus accumbens – Orbitofrontale cortex – Parahippocampale gyrus