- Introduction
- De genetische basis van hybride breuk in rijst
- Moleculaire Mechanismen die ten grondslag liggen aan hybride breuk
- Hybrid Breakdown in Conventional Rice Breeding
- Hybride breuk in Genomics-Assited Breeding
- Discussie
- Data Availability Statement
- Author Contributions
- Conflict of Interest
- Acknowledgments
Introduction
Breeders en onderzoekers zijn geïnteresseerd in hybridisatie en introgressie tussen divergerende genotypes als de potentiële motor die leidt tot de ecologische divergentie van de nakomelingen (Rieseberg et al., 2007; Arnold et al., 2012). Omgekeerd kan hybridisatie soms gepaard gaan met een verminderde hybride fitness, zoals zwakte en/of steriliteit in F1 en latere generaties, zelfs in gevallen van hybridisatie tussen leden van dezelfde soort.
Verminderde hybride levensvatbaarheid en/of vruchtbaarheid segregerend in F2 of latere generaties worden aangeduid als hybride afbraak (HB), waarbij recessieve allelen noodzakelijkerwijs geassocieerd zijn. Deze voortplantingsbarrière is lange tijd waargenomen bij zowel planten als dieren (Dobzhansky, 1970; Grant, 1971). Uit vele genetische analyses van deze barrière is gebleken dat zij tot stand komt door een complementair effect tussen en/of tussen loci met gedifferentieerde allelen, gewoonlijk de Bateson-Dobzhansky-Muller (BDM) incompatibiliteit genoemd (Rieseberg en Willis, 2007). HB gaat noodzakelijkerwijs gepaard met intrinsieke postzygote reproductieve barrières, zoals hybride onkwetsbaarheid (met inbegrip van zwakte, necrose en chlorose) en hybride steriliteit (in de mannelijke, vrouwelijke of beide gameten); daarom is het mogelijk dat sommige onderzoekers HB niet onderscheiden van onkwetsbaarheid en steriliteit in de F1-produktie. Niettemin ben ik van mening dat HB een handige classificatie is voor reproductieve barrières, omdat het hun onderliggende genetische basis impliceert (d.w.z. de betrokkenheid van recessieve allelen).
Recente studies met behulp van de Arabidopsis-modelplantensoort hebben een beter inzicht verschaft in HB met betrekking tot de moleculaire mechanismen ervan (Vaid en Laitinen, 2019). Omgekeerd, hoewel rijst (Oryza species) een modelgewas is, blijft het huidige begrip van de genetische basis van HB beperkt in deze soort, waarschijnlijk omdat HB geen reproductieve barrière is in F1 hybriden, waarin een hogere graanproductie wordt verwacht in vergelijking met de ouderlijke inteeltlijnen, en waarschijnlijk omdat inferieure planten die segregeren in F2 en latere generaties gemakkelijk kunnen worden uitgeselecteerd op basis van het fenotype uit de kweekpopulatie. De moeilijkheid in genetische kartering veroorzaakt door recessieve overerving kan ook aan deze beperking ten grondslag liggen.
Hier schets ik HB in rijstkruisingen onder verwijzing naar informatie uit Arabidopsis-studies, en bespreek ik hoe HB in de rijstveredeling wordt behandeld.
De genetische basis van hybride breuk in rijst
Men gaat ervan uit dat gecultiveerde rijst, O. sativa japonica en indica, een soortencomplex vormt met hun vermeende stamvader (O. rufipogon) en wilde soorten; echter, meerdere reproductieve barrières, waaronder HB, worden waargenomen in kruisingen tussen hen (Oka, 1988; Vaughan et al.,
Voor zover ik weet, was Dr. Oka de eerste die de genetische basis beschreef van HB in intersubspecifieke kruisingen tussen japonica en indica, die werd verklaard door twee complementaire genen (Oka, 1957). De verantwoordelijke genen werden echter niet in kaart gebracht op chromosomen, omdat moleculaire markers toen nog niet beschikbaar waren.
In rijsthybriden is HB vaak beschreven in intersubspecifieke (O. sativa ssp. japonica × ssp. indica) en interspecifieke (O. sativa × O. nivara en O. sativa × O. glumaepatula) kruisingen (Oka, 1957; Sato en Morishima, 1988; Wu et al, 1995; Li et al., 1997; Fukuoka et al., 1998, 2005; Kubo en Yoshimura, 2002, 2005; Matsubara et al., 2007a,b, 2015; Yamamoto et al., 2007; Ichitani et al., 2012;voor intersubspecifieke kruisingen; Sobrizal et al., 2001; Miura et al., 2008 voor interspecifieke kruisingen) (Tabel 1). De meeste van deze HB gevallen werden veroorzaakt door twee-locus BDM incompatibiliteit en recessieve allelen, waarbij 1/16 van de F2 nakomelingen die homozygoot waren voor recessieve allelen op beide loci het HB fenotype vertoonden; echter, 4/16 van de nakomelingen die heterozygoot waren op slechts één locus vertoonden het HB fenotype, afhankelijk van de kruiscombinatie (Tabel 1 en Figuur 1A). In de terugkruisingshybriden was één locus al gefixeerd met de allelen van de terugkerende ouder; daarom vertoonde 1/4 van de BCnF2 nakomelingen het HB fenotype (Tabel 1 en Figuur 1B).

Tabel 1. Genetische basis van de hybride afbraak gemeld in rijst kruisingen.
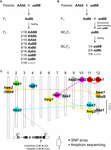
Figuur 1. De genetische basis van hybride afbraak (HB). (A) Schematische voorstelling van het HB-genotype dat segregeert in de F2-nakomelingen. Wanneer de ouderlijke genotypen AAbb en aaBB zijn, vertoont 1/16 van de F2-genotypen zonder dominant allel, dat wordt aangegeven met grijs teken (d.w.z. aabb), verminderde levensvatbaarheid en/of vruchtbaarheid, hoewel de andere genotypen normaal zijn. In sommige rijstkruisingen vertonen de F2-gen nakomelingen met slechts één dominant allel (d.w.z. Aabb, aaBb) ook verminderde levensvatbaarheid en/of vruchtbaarheid. (B) Schematische voorstelling van het HB genotype dat segregeert in de BCnF2 nakomelingen. Verder vertoont 1/4 van de BCnF2 nakomelingen verminderde levensvatbaarheid en/of vruchtbaarheid. (C) Chromosomale locatie van genen die ten grondslag liggen aan HB in rijstkruisingen. De genen zijn ruwweg in kaart gebracht op basis van de resultaten van genkartering in elke studie. Een set van complementaire genen zijn verbonden door de stippellijn. Complementaire genen met hbd1 en hwf1 zijn niet gerapporteerd. Het voorbeeld van het functionele nucleotidenpolymorfisme dat HB veroorzaakt, gepubliceerd door Yamamoto et al. (2010) wordt getoond, en deze informatie zal ons in staat stellen om een onderzoek uit te voeren door middel van SNP array of amplicon sequencing.
Het HB fenotype in rijsthybriden varieert naargelang de kruiscombinatie. Bijvoorbeeld, het hbd2/hbd2 hbd3/hbd3 genotype vertoont zwakte, maar geen duidelijke zaadsteriliteit. Het hbd4/hbd4 hbd5/hbd5 genotype daarentegen vertoont zowel zwakte als zaadsteriliteit (Matsubara et al., 2007a, 2015; Yamamoto et al., 2007; Tabel 1). Er zijn ook ingewikkelde gevallen waarin HB die zowel zwakte als zaadsteriliteit vertonen, worden waargenomen (hwe1/hwe1 hwe2/hwe2 genotype) of HB die alleen zaadsteriliteit vertonen (hsa1/hsa1 hsa2/hsa2 hsa3/hsa3), ondanks dezelfde kruiscombinatie, d.w.z. Asominori × IR24 kruising (Kubo en Yoshimura, 2002, 2005; Tabel 1). Aangezien deze HB’s onafhankelijke genetische bases hebben, zouden ten minste 5/64 van de F2 nakomelingen het HB fenotype in deze kruising vertonen.
Tot op heden zijn verschillende reeksen loci die verantwoordelijk zijn voor HB in kaart gebracht in bepaalde genomische regio’s met behulp van DNA-merkers in kruisingen tussen de japonica en indica rijstvariëteiten (Chrs 7 en 10, Fukuoka et al, 1998; de genen 1 en 12, Wu et al., 1995; Kubo en Yoshimura, 2002; Matsubara et al., 2015; de genen 6 en 11, Fukuoka et al., 2005; de genen 8, 9, en 12, Kubo en Yoshimura, 2005; de genen 2 en 11, Matsubara et al., 2007a; Yamamoto et al., 2007, 2010; de genen 11 en 12, Ichitani et al., 2012; Tabel 1 en Figuur 1C). De resultaten van deze studies toonden aan dat loci die aan de basis liggen van rijst HB gedeeld worden in sommige kruisingen, maar verschillen in andere kruisingen, wat suggereert dat ze gediversifieerd zijn in de genomen van rijst. Bijvoorbeeld, in sommige gevallen deelt het HB-allel gedragen door O. nivara, een soort die nauw verwant is aan incica-variëteiten, dezelfde locus met een indica-variëteit; maar het HB-allel gedragen door O. glumaepatula, een soort die nauw verwant is aan gecultiveerde variëteiten, deelt de locus met geen enkele andere variëteit (Tabel 1 en Figuur 1C). Opgemerkt moet worden dat de hierboven beschreven HB gevallen veroorzaakt worden door een set genen met grote effecten; er zijn echter ook gevallen die veroorzaakt worden door een set genen met kleine effecten, bijv. licht verminderde zaadvruchtbaarheid, die niet over het hoofd gezien kunnen worden in de rijstveredeling, die nog niet ontdekt zijn.
Moleculaire Mechanismen die ten grondslag liggen aan hybride breuk
Autoimmuunrespons: Bomblies et al. (2007) meldden voor het eerst dat de auto-immuunrespons die een NB-LRR-gen tegen ziekten of een R-gen veroorzaakt, geassocieerd is met Arabidopsis HB in intraspecifieke kruisingen, hoewel deze auto-immuunrespons vooral tot uiting komt als necrose in F1-planten, en HB in F2-nakomelingen geconditioneerd leek te zijn door de temperatuur. In Arabidopsis hybriden zijn verschillende lijnen van bewijs van HB veroorzaakt door de auto-immuunrespons beschreven (Alcázar et al., 2009, 2010). De betrokkenheid van NB-LRR genen bij HB suggereert dat meerdere genomische regio’s geassocieerd kunnen worden met de Arabidopsis HB; in feite ondersteunt het uitgebreide onderzoek uitgevoerd door Chae et al. (2014) dit idee.
Ook in rijsthybriden is HB veroorzaakt door een auto-immuunrespons gerapporteerd in een japonica × indica kruising (Yamamoto et al., 2010). In dit geval treedt HB op in F2 planten wanneer het hybride afbraak 2 (hbd2) gen, dat codeert voor caseïne kinase I en wordt gedragen door de indica variëteit, wordt gecombineerd met een cluster van R genen gedragen door de japonica variëteit. Aangezien deze oorzakelijke allelen zich waarschijnlijk als gedeeltelijk recessieve allelen gedragen, kunnen de genproducten hierbij betrokken zijn (Matsubara et al., 2007a; Yamamoto et al., 2007, 2010).
Reciprocal silencing of duplicated genes: dit begrip werd voor het eerst theoretisch voorgesteld als een genetische basis voor hybride incompatibiliteit (Werth en Windham, 1991; Lynch en Force, 2000). In het model verliest de ene populatie functie op de ene locus en behoudt die op de andere, terwijl de andere populatie het tegenovergestelde effect ondervindt. Dientengevolge heeft 1/16 van de F2 zygoten geen functionele genen (Lynch en Force, 2000). De HB in Arabidopsis hybriden beschreven door Bikard et al. (2009) en Vlad et al. (2010) is een goed voorbeeld van dit fenomeen. In dit scenario zijn ook gevallen gemeld waarbij epigenetische silencing een rol speelt (Agorio et al., 2017; Blevins et al., 2017).
Omdat rijst zowel hele-genoom- als segmentale duplicatie heeft meegemaakt (Wang et al., 2007; Guo et al., 2019), zou wederkerige silencing de overhand kunnen hebben als oorzaak van hybride incompatibiliteit. Tot op heden is er echter geen bewijs voor HB veroorzaakt door wederzijdse silencing van gedupliceerde genen in rijst.
Onafhankelijk van de auto-immuunrespons en wederzijdse silencing, is een interactie tussen een DEAD-box RNA helicase 18 en een ander gen (MORPHEUS MOLECULE 1 als een waarschijnlijke kandidaat) geïdentificeerd die HB veroorzaken in Arabidopsis-hybriden (Plötner et al., 2017; Vaid et al., 2020). Interessant is dat het HB fenotype vermindert tussen de F3 en F4 generaties, wat epigenetische regulatie van genexpressie impliceert.
In rijsthybriden, onder de drie hybride steriliteit-a (hsa) loci, toonden Kubo et al. (2016) onlangs aan dat de hsa1 locus uit twee genen bestaat, en dat deze genen coderen voor respectievelijk een DUF1618 eiwit en een ongekarakteriseerd eiwit met enige gelijkenis met een nucleotide-bindend eiwit. De moleculaire kenmerken van de overige complementaire genen, hsa2 en hsa3, zijn niet gerapporteerd.
Hybrid Breakdown in Conventional Rice Breeding
Tientallen jaren lang werd de bulk-populatie methode op grote schaal toegepast in de conventionele rijstveredeling. Bij deze methode wordt na kruising de F4- of F5-populatie grootgebracht door zelfbevruchting in bulk (in plaats van door de afstammingsmethode met één zaadje) zonder kunstmatige selectie, maar de populatie van de eerste generatie wordt onderworpen aan natuurlijke selectie en selectie op levensvatbaarheid (Allard, 1960; Ikehashi en Fujimaki, 1980 voor details). Bij de bulk-populatiemethode zullen naar verwachting veel zwakke en/of steriele genotypen uit de populatie zijn geëlimineerd voordat een populatie van een gevorderde generatie is gevormd; daarom kan deze methode beperkte informatie over HB opleveren.
Voor de introgressie of accumulatie van gewenste eigenschappen van de donor(en) naar een variëteit hebben rijstveredelaars vaak terugkruisingen of meervoudige ouderkruisingen uitgevoerd, gevolgd door de bulk-populatiemethode. Empirisch is reeds lang bekend dat deze kruisingsmethoden het verlies van selectiekandidaten door voortplantingsbarrières kunnen beperken, omdat deze methoden vaak de segregatie van storende combinaties van allelen die geassocieerd worden met HB in een hybride nageslacht verminderen. De mitigatie van reproductieve barrières door deze kruisingsmethoden is echter onvermijdelijk afhankelijk van het HB-genotype van de ouders.
Hybride breuk in Genomics-Assited Breeding
De publicatie van genoomsequenties van referentiegewassen en de ontwikkeling van next-generation sequencing-technologieën hebben de vooruitgang van de moleculaire veredeling van gewassen versneld (Kole et al., 2015). In deze context is genomics-assisted breeding, zoals genoomselectie op basis van genoombrede DNA-merkers, overwogen in de gewasveredeling (Spindel en Iwata, 2018).
In genomics-assisted breeding van zelfbestoven gewassen, worden populaties van geavanceerde generaties, zoals recombinant inteeltlijnen, vaak gebruikt als referentiepopulaties, waarvan genoombrede genotype- en fenotypegegevens worden verkregen. Vervolgens wordt selectie op basis van uitsluitend markergenotypes uitgevoerd in de nakomelingen van vroege generaties (bv. F2). Bij dit selectieschema moet worden opgemerkt dat de referentiepopulatie gewoonlijk geen informatie over het genomische gebied voor HB verschaft, omdat zwakke en/of steriele nakomelingen in de vroege generaties na kruising moeten zijn geëlimineerd. Als een dergelijk selectieschema wordt toegepast, zullen we dus een aantal belangrijke selectiekandidaten moeten laten vallen. Een andere mogelijkheid is dat we ongewenste kandidaten selecteren, zodat HB in latere generaties duidelijk wordt.
Discussie
Ondanks het beperkte aantal studies over dit onderwerp, lijkt het erop dat de loci die ten grondslag liggen aan HB in rijstkruisingen veeleer gediversifieerd dan gedeeld zijn (tabel 1 en figuur 1). Zoals hierboven beschreven, zou de informatie over de verdeling van HB-geassocieerde allelen onder kruisingsouders een eerste vereiste moeten zijn voor de rijstveredeling, in het bijzonder voor het maximaliseren van de effectiviteit van veredeling met behulp van genomica. Daarom is de aanvullende opsporing van HB in rijstkruisingen en het in kaart brengen van verantwoordelijke genen in het rijstgenoom nodig. In Arabidopsis hybriden zijn dergelijke gegevens over de R-genen en hun interagerende genen uitgebreid in kaart gebracht (Alcázar et al., 2010, 2014; Chae et al., 2014). De rapportage van het hbd2 gen door Yamamoto et al. (2010) in een rijsthybride is een goed voorbeeld van een uitgebreid onderzoek uitgevoerd met SNP arrays of amplicon sequencing (Figuur 1C). De hbd3 en hsa1 genen zijn ook kandidaten voor dit type onderzoek. Uiteindelijk zal de ontwikkeling van criteria die ons in staat stellen HB te voorspellen op basis van de genomische informatie van de ouderlijnen, een belangrijk instrument vormen voor veredeling met behulp van genomics. Zelfs als een oorzakelijke factor niet is geïdentificeerd als een enkel gen, zullen nauw verbonden merkers, zoals single-nucleotide polymorfismen, effectief zijn voor het classificeren van HB-geassocieerde haplotypes, omdat het linkage disequilibrium van gecultiveerde rijst is geraamd op 100-200 kb, terwijl dat van wilde soorten (zoals O. rufipogon) kan zich uitstrekken over enkele tientallen kb (Mather et al., 2007; McNally et al., 2009; Huang et al., 2010). Met deze informatie kan een efficiëntere en effectievere kruiscombinatie worden ontworpen.
Bulked segregant analyse gevolgd door next-generation sequencing kan nuttig zijn voor het in kaart brengen van HB-geassocieerde loci, evenals conventionele linkage mapping (zoals kwantitatieve trait locus analyse), omdat HB-segreganten vaak onderscheidbare kenmerken vertonen (over eigenschappen zoals planthoogte, kiemgetal, en vruchtbaarheid) van normale groei segreganten in elke kruispopulatie. Het verzamelen van beeldgegevens met behulp van een digitale camera en het laden met een drone kan ook een belangrijke rol spelen bij de fenotypering in het laboratorium en op het rijstveld. Deze inspanningen zullen niet alleen waardevolle informatie opleveren voor rijstveredelaars, maar ook voor evolutiebiologen.
Verder, als een HB-geassocieerd allel wordt geïdentificeerd, kan genverstoring door ionenbundels of genoombewerking helpen HB te overwinnen, zoals geïllustreerd door F1 hybride steriliteit in rijst (Koide et al., 2018; Xie et al., 2019).
Data Availability Statement
Alle datasets die in deze studie worden gepresenteerd, zijn opgenomen in het artikel/aanvullend materiaal.
Author Contributions
De auteur bevestigt de enige bijdrager van dit werk te zijn en heeft het goedgekeurd voor publicatie.
Conflict of Interest
De auteurs verklaren dat het onderzoek werd uitgevoerd in afwezigheid van enige commerciële of financiële relaties die zouden kunnen worden opgevat als een potentieel belangenconflict.
Acknowledgments
Ik zou dit artikel willen opdragen aan de nagedachtenis van Dr. Darshan Brar. Hij was een eminent rijstveredelaar die met grote voortvarendheid nuttige genen, gedragen door wilde soorten, introduceerde in cultivars voorbij de reproductieve barrières.
Allard, R. W. (1960). Beginselen van de plantenveredeling. New York, NY: John Wily and Sons, Inc.
Google Scholar
Dobzhansky, T. (1970). Genetica van het evolutionaire proces. New York, NY: Columbia University Press.
Google Scholar
Grant, V. (1971). Plant Speciation. New York, NY: Columbia University Press.
Google Scholar
Ikehashi, H., and Fujimaki, H. (1980). “Modified bulk population method for rice breeding,” in Proceedings of the International Rice Research Conference on Innovative approaches to rice breeding, (Los Baños, PA: International Rice Research Institute), 163-182.
Google Scholar
Oka, H.-I. (1988). Oorsprong van gecultiveerde rijst. Tokio: JSSP.
Google Scholar