Serum albumine
Structure
Serum albumine is het meest voorkomende eiwit in het bloedplasma van alle gewervelde dieren, waarbij de concentratie in menselijk serum 35-50 mg/mL bedraagt (Peters, 1996). Humaan serumalbumine (HSA) heeft een moleculaire massa van 66 348 Da en is samengesteld uit drie homologe domeinen, genummerd I, II en III (figuur 1) (He en Carter, 1992; Peters, 1996; Sugio et al., 1999). Elk domein is gegroepeerd in subdomeinen A en B die gemeenschappelijke structurele motieven bezitten. De twee belangrijkste regio’s die verantwoordelijk zijn voor ligand-binding aan HSA staan bekend als Sudlow’s Site I en II, respectievelijk gelegen in subdomein IIA en IIIA (figuur 1) (Sudlow et al., 1976; Peters, 1996). Albumine wordt gecodeerd door één enkel gen, dat op een co-dominante manier tot expressie komt, waarbij beide allelen worden getranscribeerd en vertaald (Hawkins en Dugaiczyk, 1982; Peters, 1996). Het menselijke albumine-gen bevindt zich op de lange arm van chromosoom 4 op positie q13.3.
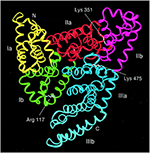
Figuur 1. Structuur van humaan serumalbumine bestaande uit drie domeinen, elk gegroepeerd in subdomeinen A en B (Subdomein Ia, geel; Ib, groen; IIa, rood; IIb, magenta; IIIa, blauw; en IIIb, cyaan). Sugio et al. (1999) met toestemming van Oxford University Press.
Functie
Albumine wordt voornamelijk door de lever gesynthetiseerd, waarbij de menselijke lever ~13,9 g HSA per dag produceert (Peters, 1996). HSA heeft een geschatte halfwaardetijd van 19 dagen en wordt effectiever afgebroken als het gedenatureerd of structureel veranderd is (Peters, 1996). Albumine heeft een verscheidenheid aan belangrijke functies en is verantwoordelijk voor 80% van de colloïdale osmotische druk van bloed (Peters, 1996). Belangrijk is dat albumine in staat is verschillende endogene moleculen te binden, waaronder lange-keten vetzuren, steroïden, L-tryptofaan, enz. (Kragh-Hansen, 1981; Peters, 1996; Evans, 2002). Bovendien is albumine ook betrokken bij het transport van ionen in de circulatie, waaronder koper, zink, calcium, enz. (Peters, 1996).
Daarnaast is dit vitale eiwit in staat om exogene verbindingen en geneesmiddelen te binden, zoals warfarine, ibuprofen, chloorpromazine en naproxen, waarbij de affiniteit van hun binding hun activiteit en halfwaardetijd aanzienlijk beïnvloedt (Kragh-Hansen, 1981; Peters, 1996; Evans, 2002). Bovendien fungeert albumine ook als drager van toxisch afval, het bindt bilirubine, het product van de afbraak van heem, om het naar de lever af te voeren voor uitscheiding door de lever (Peters, 1996). Interessant is dat albumine ook wordt verondersteld te werken als een anti-oxidant vanwege zijn vermogen om: (1) gebonden stoffen te beschermen tegen peroxidatieve schade (bijv. vetzuren en lipoproteïnen); en (2) vrij koper te binden, waardoor de redoxactiviteit en de productie van vrije radicalen beperkt worden (Peters, 1996; Evans, 2002). Tenslotte is albumine een bron van thiolen die gretige reactieve zuurstof- en stikstofspecies opruimers zijn (Peters, 1996; Evans, 2002).
Distributie
Het is interessant dat albumine overwegend aanwezig is in de extravasculaire ruimte (~242 g) in plaats van de intravasculaire ruimte (~118 g) (Peters, 1996; Evans, 2002). In feite is het eiwit overwegend aanwezig in extracellulaire locaties zoals huid, darm, spieren, andere vloeistoffen (d.w.z. cerebrospinaal, pleuraal, enz.) en afscheidingen (d.w.z. zweet, tranen en melk) (Peters, 1996). Intracellulair zijn echter zeer lage concentraties albumine aanwezig (Peters, 1996). Albumine keert terug van de extravasculaire ruimte naar de circulatie via het lymfestelsel, en maakt ~28 “reizen” in en uit het lymfestelsel tijdens zijn leven (Peters, 1996; Evans, 2002).
Na afscheiding uit hepatocyten komt albumine in de circulatie en transloceert naar de extracellulaire ruimte door de poriën van sinusoïdaal of venestervormig endotheel in bepaalde organen, zoals de lever, pancreas, dunne darm en beenmerg (Peters, 1996). In organen waar een continu endotheel overheerst, wordt nu echter aangenomen dat albumine het endotheel kan passeren via actieve transcytotische mechanismen, waaronder receptormedieerde mechanismen (b.v, albondine; zie het hoofdstuk “Cellulaire albumine-bindende eiwitten”).
Accumulatie van albumine in het tumorinterstitium
Vaste tumoren bezitten gewoonlijk een onrijpe, zeer permeabele vasculatuur die wordt beïnvloed door vasculaire permeabiliteitsverhogende factoren (b.v, stikstofmonoxide) (Carmeliet en Jain, 2000; Maeda et al., 2000; Greish, 2007; van der Veldt et al., 2008). Desondanks is er over het algemeen onvoldoende lymfedrainage (Carmeliet en Jain, 2000; Maeda et al., 2000; Greish, 2007). Dit leidt vervolgens tot een ophoping van macromoleculen (>40 kDa) in het tumorinterstitium, en dit staat bekend als het versterkte permeatie- en retentie-effect (figuur 2) (Maeda et al., 2000; Greish, 2007). Van belang is dat Matsumura en Maeda (1986) aantoonden dat een intraveneus geïnjecteerd Evans blauw-albumine complex zich ophoopte in sarcoma 180 tumoren van ddY muizen. De retentie van albumine in tumoren is sindsdien waargenomen in verschillende experimentele vaste tumoren (b.v. sarcoom, ovariumcarcinoom, Novikof hepatoma, enz.) met gebruikmaking van radioactief gelabeld of met kleurstof gecomplexeerd serumalbumine (Peterson en Appelgren, 1973; Sinn et al, 1990; Andersson et al., 1991; Schilling et al., 1992; Stehle et al., 1997; Wunder et al., 1997).
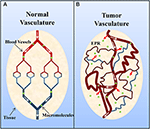
Figuur 2. Schematische voorstelling van (A) normale en (B) tumor vasculatuur. Normaal weefsel bestaat uit volgroeide, georganiseerde bloedvaten, terwijl tumorweefsel bestaat uit onrijpe, lekkende en kronkelige vaten. De veranderde organisatie van tumor vasculatuur en ongeorganiseerd lymfatisch netwerk resulteert in vasculaire lekkage en de ophoping van macromoleculen (>40 kDa) in het interstitium en staat bekend als het enhanced permeation and retention (EPR) effect. Overgenomen met toestemming van Macmillan Publishers Ltd: Nature Medicine (Jain, 2001), copyright (2001).
Verder is in een aantal studies voorgesteld dat tumoren een plaats zijn van albumine katabolisme (Hradec, 1958; Andersson et al., 1991; Schilling et al., 1992; Stehle et al., 1997). Bijvoorbeeld, in een sarcomamodel van de muis (C57/RL6J), geïnjecteerd met 3H-raffinose gelabeld albumine, werden ten minste 2-3-voudig hogere niveaus van 3H waargenomen in de lysosomen van tumoren in vergelijking met lysosomen van normaal weefsel (Andersson et al., 1991). Verder hebben studies aangetoond dat albumine een kortere halfwaardetijd heeft en een hogere turnover in tumordragende muizen, ondanks een compenserende toename in de leveralbuminesynthese, vergeleken met niet-tumordragende muizen (Hradec, 1958). Vandaar dat er is gesuggereerd dat tumoren albumine gebruiken als energiebron, door het afbreken van albumine in de samenstellende aminozuren in lysosomen, die vervolgens door kankercellen worden gebruikt voor hun versnelde groei (Stehle et al., 1997). Bovendien hebben studies gesuggereerd dat de hypoalbuminiemie die duidelijk is bij kankerpatiënten het resultaat is van albumine katabolisme door de tumor (Stehle et al., 1997).
Niettemin hebben sommige van deze eerdere studies te lijden onder verschillende experimentele beperkingen. Het is bijvoorbeeld moeilijk om zuivere lysosomale fracties te verkrijgen en daarom is het nodig om deze studies te reproduceren en de zuiverheid van fracties te testen met behulp van goed gevestigde membraan- en organelmarkers (Graham, 2002; Yamagishi et al., 2013). Bovendien kunnen verschillende andere in vivo factoren de afbraak en het katabolisme van albumine beïnvloeden (bv. niveaus van corticosteroïden) (Peters, 1996). Daarom is een duidelijke relatie niet vastgesteld en zijn aanvullende in vivo studies nodig om de intracellulaire distributie en het katabolisme van albumine door tumoren te ondersteunen.
Meer recent hebben Commisso et al. waargenomen dat kankercellen met endogene oncogene Ras-mutaties verhoogde niveaus van macropinocytose in vitro en in vivo hebben (Commisso et al., 2013). Bovendien werd aangetoond dat FITC-gelabeld albumine werd geïnternaliseerd via macropinocytose en vervolgens resulteerde in verhoogde niveaus van glutamaat en α-ketoglutaraat in oncogene Ras-getransformeerde cellen (Commisso et al., 2013). Interessant is dat de afname in proliferatie van oncogene Ras-expresserende cellen na glutamine deprivatie bleek te worden hersteld door extracellulaire albumine suppletie (Commisso et al., 2013). Deze bevindingen suggereren dat macropinocytose van albumine voedingsstoffen levert om de proliferatie van kankercellen in stand te houden (Commisso et al., 2013).
Cellulaire albuminebindende eiwitten
Gezien het belang van albumine, is een aantal putatieve albuminebindende eiwitten en receptoren geïdentificeerd in verschillende weefsels en cellijnen (tabel 1), waaronder de nier (Zhai et al., 2000; Amsellem et al., 2010), endotheel (Schnitzer en Bravo, 1993), fibroblasten (Porter et al., 1995), en tumorceloppervlakken (Fritzsche et al., 2004). In het bijzonder zijn er zeven membraan-geassocieerde albumine-bindende eiwitten ontdekt, namelijk: albondine/glycoproteïne 60 (gp60) (Schnitzer et al., 1988), glycoproteïne 18 (gp18) (Ghinea et al., 1988), glycoproteïne 30 (gp30) (Ghinea et al, 1988), de neonatale Fc-receptor (FcRn) (Roopenian en Akilesh, 2007), heterogene nucleaire ribonucleoproteïnen (hnRNPs) (Fritzsche et al., 2004), calreticulin (Fritzsche et al., 2004), cubilin (Zhai et al., 2000; Amsellem et al., 2010), en megalin (Zhai et al., 2000; Amsellem et al., 2010). Bovendien is een afgescheiden albuminebindend eiwit geïdentificeerd dat bekend staat als secreted protein, acidic and rich in cysteine (SPARC) (Schnitzer en Oh, 1992). Gezien hun belang bij de opname van albumine door cellen, wordt elk van deze eiwitten hieronder in detail beschreven.

Tabel 1. Localization of albumin-binding proteins and receptors.
Albondin/gp60
Albondin (gp60) is een 60 kDa glycoproteïne dat fungeert als een albumine-receptor die wijd verspreid is, maar selectief tot expressie komt op het plasmamembraan van continu endotheel (behalve in de hersenen), waar het de capillaire permeabiliteit verhoogt (Ghinea et al., 1988, 1989; Schnitzer et al., 1988; Schnitzer, 1992; Schnitzer en Oh, 1994; Tiruppathi et al., 1996). Albondine bindt niet alleen specifiek natief albumine, maar vergemakkelijkt ook de internalisatie en daaropvolgende transcytose (Milici et al., 1987; Schnitzer, 1992; Schnitzer and Oh, 1994; Tiruppathi et al., 1996).
Voorgesteld is dat ~50% van albumine het capillair verlaat via albondine, waarbij de rest deze barrière passeert via intercellulaire juncties en/of vloeistof-fase mechanismen (Schnitzer, 1993; Schnitzer and Oh, 1994). Bovendien is aangetoond dat de internalisatie van albondine plaatsvindt via een caveolin-afhankelijk endocytotisch proces dat resulteert in transcytose en niet lijkt in te gaan op het degradatieve endosoom-lysosoom systeem (Schnitzer, 1993; Schnitzer en Bravo, 1993; Schnitzer et al., 1995; Tiruppathi et al., 1997; Iancu et al., 2011).
gp18 en gp30
Zowel gp18 als gp30 binden zich gretig aan conformationeel gemodificeerd albumine (d.w.z, met goud gelabeld albumine, formaldehyde- of maleïnezuuranhydridebehandeld albumine) en hebben geen preferentiële interactie met natief albumine, vergelijkbaar met andere bekende scavenger receptoren (Ghinea et al., 1989; Schnitzer en Oh, 1992, 1994; Schnitzer et al., 1992; Schnitzer en Bravo, 1993). In tegenstelling tot albondine, worden gp18 en gp30 gevonden op een verscheidenheid van cellen, zoals macrofagen en fibroblasten, en een reeks van endotheelcellen (Schnitzer et al., 1992). Bovendien werd vastgesteld dat gp18 tot expressie komt in menselijke MDA-MB-453 borstkankercellen (Wang et al., 1994). Deze scavenger receptoren binden en leiden gemodificeerde albumines voor degradatie, misschien als onderdeel van eiwitkatabolisme of als een beschermende route om veranderde, oude, beschadigde of potentieel schadelijke albumines te verwijderen (Schnitzer, 1993; Schnitzer en Bravo, 1993). Albumine kan worden gemodificeerd door oxidatie, niet-enzymatische glycatie, maleylering, etc. als gevolg van normale veroudering of als een beschermende of pathologische reactie (Schnitzer, 1993; Peters, 1996). Gedenatureerd of gemodificeerd albumine wordt sneller en efficiënter afgebroken dan natief albumine, wat suggereert dat deze veranderingen albuminemoleculen selecteren voor afbraak (Peters, 1996).
SPARC
SPARC is ook bekend als osteonectine en BM-40 en wordt door verschillende celtypen uitgescheiden (Brekken en Sage, 2001). Interessant is dat SPARC sterk tot expressie komt in maligne cellen en stromale cellen die geassocieerd worden met neoplasie (Porter et al., 1995; Podhajcer et al., 2008). SPARC bezit albumine-bindende eigenschappen en interageert specifiek met natief albumine op een vergelijkbare manier als albondine, maar verschillend van 18 tot gp30 die conformationeel gewijzigd albumine binden (Schnitzer en Oh, 1992). Specifiek herkennen anti-SPARC antilichamen ook albondine, maar niet 18 of gp30, wat suggereert dat SPARC en albondine een natief albumine-bindend domein delen (Schnitzer en Oh, 1992). Er is echter geen bewijs dat SPARC de opname van albumine in tumoren medieert. Er is gesteld dat het vermogen van SPARC om albumine te binden in het tumorinterstitium de accumulatie van albumine-gebonden geneesmiddelen in de tumorruimte bevordert (Desai et al., 2008, 2009). Bovendien toonde een voorlopige klinische studie aan dat SPARC-expressie correleerde met de respons op behandeling met paclitaxel-geladen nanodeeltjes van albumine (nab-paclitaxel of Abraxane®), waarbij SPARC-positieve patiënten een betere klinische uitkomst hadden (Desai et al., 2009). Tegenstrijdige gegevens in een KPfC-muismodel hebben deze hypothese echter in twijfel getrokken, aangezien SPARC-deficiëntie de intra-tumorale concentraties van Abraxane® niet veranderde (Neesse et al., 2014). Bijgevolg zijn verdere studies nodig om deze hypothese te valideren, met inbegrip van grotere klinische studies met een groter aantal patiënten. Momenteel wordt in een fase III-studie (NCT00785291), door het National Cancer Institute, geëvalueerd of SPARC-expressie in serum de respons van patiënten op Abraxane® voorspelt, en dit kan ons inzicht in de rol van SPARC in albumine-accumulatie door tumoren bevorderen.
hnRNPs en Calreticulin
Vijf verschillende albumine-bindende eiwitten zijn geïdentificeerd uit plasmamembranen van menselijke kankercellijnen (d.w.z., CCRF-CEM T-cell leukemie, MV3 melanoom en MCF7 borstcarcinoom) (Fritzsche et al., 2004). Vier van deze eiwitten werden geïdentificeerd als leden van de hnRNP-familie, waaronder hnRNP A2/B1, hnRNP C1, hnRNP A1 en hnRNP A3, en het vijfde eiwit bleek calreticulin te zijn (Fritzsche et al., 2004). Calreticuline werd voor het eerst beschreven als een endoplasmatisch reticulum chaperon en calcium signaleringseiwit, maar sindsdien is aangetoond dat het betrokken is bij verschillende cellulaire functies, waaronder celadhesie, modulatie van bloedplaatjes-collageen interacties (wondgenezing) en apoptose (Mendlovic en Conconi, 2010). De functies van de hnRNP-familie zijn niet goed gekarakteriseerd. De meeste leden van de hnRNP-familie zijn echter beschreven als nucleaire RNA-bindende eiwitten die betrokken zijn bij de verwerking van pre-mRNA, zoals RNA-splicing, export en stabiliteit (Chaudhury et al., 2010). Interessant is dat hnRNPs zijn voorgesteld om een rol te spelen in carcinogenese, waar hun overexpressie fungeert als biomarkers voor de vroege detectie van tumoren (Han et al., 2013). De betekenis van deze bevindingen is momenteel onduidelijk en het moet nog worden vastgesteld of deze eiwitten betrokken zijn bij albumine-gemedieerde opname.
FcRn
FcRn komt tot expressie in meerdere celtypen en weefsels, waaronder antigeen-presenterende cellen, vasculair endotheel, darm, longen, nieren en de bloed-hersenbarrière (BBB) (d.w.z, endotheel en plexus choroideus) (Roopenian en Akilesh, 2007). Deze receptor beschermt albumine en IgG tegen afbraak door beide eiwitten alleen bij een lage pH (pH < 6,5) in zure endosomen met hoge affiniteit te binden, waardoor hun afbraak via de lysosomale route wordt voorkomen en ze terugkeren naar de extracellulaire ruimte (pH 7,4) (Chaudhury et al., 2003; Ober et al., 2004; Anderson et al., 2006; Andersen et al., 2012). Dit verlengt bijgevolg de halfwaardetijd van serumalbumine (Chaudhury et al., 2003; Anderson et al., 2006; Sarav et al., 2009). De rol van deze receptor bij de opname van albumine door tumoren blijft onduidelijk.
Cubilin en Megalin
Cubilin is een multi-ligand receptor die het meest bekend is vanwege zijn betrokkenheid bij de intestinale opname van de intrinsieke factor vitamine B12 complex (Seetharam et al., 1997). Bovendien is aangetoond dat cubiline betrokken is bij de endocytose en het transcellulaire transport van talrijke liganden, waaronder albumine (Birn et al., 2000). Cubiline is gelokaliseerd in absorberende darmcellen, placenta, viscerale dooierzakcellen en proximale tubuli van de nieren (Christensen en Birn, 2002). Megalin is een groot trans-membraaneiwit waarvan is aangetoond dat het ook albumine bindt (Cui et al., 1996). Dit eiwit komt op grotere schaal tot expressie dan cubiline en is aanwezig in de plexus choroideus, proximale tubuluscellen van de nieren, thyrocyten, enz. (tabel 1) (Christensen en Birn, 2002).
Interessant is dat megalin zich met hoge affiniteit aan cubilin bindt en er is gesuggereerd dat megalin als co-receptor bijdraagt tot de internalisatie van cubilin-ligandcomplexen (Moestrup et al., 1998; Christensen en Birn, 2002). Bovendien bindt cubilin ook amnionloos, een eiwit dat noodzakelijk is voor de expressie van cubilin op de celmembraan (Amsellem et al., 2010). Cubiline speelt, samen met megalin, een essentiële rol bij de opname van albumine (d.w.z. reabsorptie) door de proximale tubuli van de nieren (Zhai et al., 2000; Amsellem et al., 2010). Cubiline- en/of megalinedeficiëntie bij muizen en honden veroorzaakt aantoonbaar een afname van de opname van albumine in de proximale tubulus, wat leidt tot albuminurie (Birn et al., 2000; Amsellem et al., 2010). Bovendien lijden patiënten met het Imerslund-Gräsbeck syndroom, veroorzaakt door een mutatie in het cubilin gen, in het algemeen aan proteïnurie, wat het belang van cubilin in de proteïne renale reabsorptie aantoont (Grasbeck, 2006).
Albumin as a Drug Carrier in Oncology
Gezien het versterkte permeatie- en retentie-effect en de accumulatie van albumine in het tumorinterstitium, is de ontwikkeling van albumine als drug carrier steeds belangrijker om te overwegen in termen van de gerichte toediening van kankertherapie (Kratz, 2008, 2010). Er is ook voorgesteld dat dragers van albumine voordeel halen uit de aanwezigheid van albondine op het endotheel en SPARC in het tumorinterstitium om de accumulatie van geneesmiddelen in de tumorruimte te verhogen (Desai et al., 2009; Kratz, 2010). Er zijn verschillende geneesmiddelafgiftesystemen met albumine ontwikkeld, waaronder albumine-nanodeeltjes, geneesmiddel-albumineconjugaten, albumine-bindende geneesmiddelderivaten en prodrugs (voor reviews zie Kratz, 2008, 2010).
De ontwikkeling en marktgoedkeuring van het met paclitaxel beladen albumine-nanodeeltje, nab-paclitaxel of Abraxane®, was een belangrijke doorbraak op het gebied van de ontwikkeling van albumin dragers. Abraxane® werd voor het eerst goedgekeurd voor klinisch gebruik in de Verenigde Staten in 2005 (Kudlowitz en Muggia, 2014). Dit albumine-nanopartikel is geïndiceerd voor de behandeling van uitgezaaide borstkanker, na falen van combinatiechemotherapie (Kudlowitz en Muggia, 2014). Meer recent is Abraxane® ook beschreven voor de eerstelijnsbehandeling van patiënten met gemetastaseerd adenocarcinoom van de pancreas, in combinatie met gemcitabine, en patiënten met lokaal gevorderd of gemetastaseerd niet-kleincellig longcarcinoom, in combinatie met carboplatine (Kudlowitz en Muggia, 2014). Abraxane® heeft een grotere therapeutische index dan paclitaxel alleen en kan in hogere doses worden toegediend met minder toxiciteit en meer werkzaamheid dan de traditionele paclitaxeltherapie (Gradishar et al., 2005; Socinski et al., 2012; Iwamoto, 2013). Abraxane® wordt momenteel nog verder geëvalueerd in klinische studies voor andere tumoren, zoals blaaskanker (NCT00583349) en multipel myeloom (NCT02075021).
Daarnaast wordt ook de binding van albumine als algemene strategie voor de verbetering van de farmacokinetiek van geneesmiddelen geëvalueerd. Traditioneel wordt aangenomen dat de binding van een geneesmiddel aan albumine het niveau van vrij geneesmiddel vermindert dat beschikbaar is om de therapeutische activiteit uit te oefenen (Lancon et al., 2004; Vuignier et al., 2010). Studies hebben echter ook mechanismen aangetoond waardoor albumine het therapeutisch gebruik effectief verbetert of een snelle klaring vermindert (Dennis et al., 2002; Merlot and Richardson, 2014). Zo is aangetoond dat het experimentele antikanker thiosemicarbazon, namelijk di-2-pyridylketon 4,4-dimethyl-3-thiosemicarbazon (Dp44mT) (Merlot et al., 2013a), door kankercellen wordt geïnternaliseerd via een putatieve drager/receptor (Merlot et al., 2013b; Merlot and Richardson, 2014). Interessant is dat de opname, toxiciteit en apoptotische activiteit van Dp44mT sterk wordt versterkt in de aanwezigheid van HSA (Merlot en Richardson, 2014). Gezien het feit dat Dp44mT zich richt op lysosomen om apoptose te induceren (Lovejoy et al., 2011), en dat HSA mogelijk lysosomaal katabolisme ondergaat in tumoren (Andersson et al., 1991; Stehle et al., 1997), kan worden verondersteld dat HSA de levering van Dp44mT aan de lysosomen vergemakkelijkt, waardoor de antikankeractiviteit wordt versterkt (Merlot en Richardson, 2014). Hoewel studies het exacte mechanisme van het HSA-gestimuleerde opnameproces nog moeten identificeren, kan albumine-binding een voordeel bieden bij het genereren van tumor targeting agents en dit vereist verder intensief onderzoek.
Conclusie
Albumine is een veelzijdig en boeiend eiwit. Gezien de grote opsluiting van albumine in de vasculaire en interstitiële ruimte, is de intracellulaire distributie van albumine vele jaren slecht gekarakteriseerd gebleven. Het is mogelijk dat albumine, onder specifieke omstandigheden of tijdens cellulaire stress, wordt opgenomen door normale cellen en tumorcellen in respectievelijk lage en hoge gehaltes, als gevolg van hun metabole snelheid. De precieze rol van sommige onvolledig gekarakteriseerde albumine-bindende eiwitten (d.w.z. hnRNPs en calreticuline) bij het mediëren van de albumine-opname moet nog worden bepaald. Het zoeken en karakteriseren van albumine-bindende eiwitten, met name in kankercellen, is echter van groot belang in het licht van de ontwikkeling van albumine als een effectieve drugdrager om tumoren aan te pakken.
Conflict of Interest Statement
De auteurs verklaren dat het onderzoek werd uitgevoerd in afwezigheid van enige commerciële of financiële relaties die zouden kunnen worden opgevat als een potentieel belangenconflict.
Acknowledgments
Angelica M. Merlot ontvangt een Early Career Research Grant van de Universiteit van Sydney. Des R. Richardson is de ontvanger van een National Health and Medical Research Council (NHMRC) Senior Principal Research Fellowship en Project Grants. Danuta S. Kalinowski ontvangt een NHMRC Project Grant (1048972) en een Helen and Robert Ellis Fellowship van de Sydney Medical School Foundation van The University of Sydney.
Amsellem, S., Gburek, J., Hamard, G., Nielsen, R., Willnow, T. E., Devuyst, O., et al. (2010). Cubilin is essentieel voor albumine reabsorptie in de renale proximale tubulus. J. Am. Soc. Nephrol. 21, 1859-1867. doi: 10.1681/ASN.2010050492
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Andersen, J. T., Dalhus, B., Cameron, J., Daba, M. B., Plumridge, A., Evans, L., et al. (2012). Structuur-gebaseerde mutagenese onthult de albumine-bindende site van de neonatale Fc-receptor. Nat. Commun. 3, 610. doi: 10.1038/ncomms1607
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Anderson, C. L., Chaudhury, C., Kim, J., Bronson, C. L., Wani, M. A., and Mohanty, S. (2006). Perspectief-FcRn transporteert albumine: relevantie voor immunologie en geneeskunde. Trends Immunol. 27, 343-348. doi: 10.1016/j.it.2006.05.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Andersson, C., Iresjo, B. M., and Lundholm, K. (1991). Identification of tissue sites for increased albumin degradation in sarcoma-dragende muizen. J. Surg. Res. 50, 156-162. doi: 10.1016/0022-4804(91)90240-M
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Birn, H., Fyfe, J.C., Jacobsen, C., Mounier, F., Verroust, P.J., Orskov, H., et al. (2000). Cubilin is an albumin binding protein important for renal tubular albumin reabsorption. J. Clin. Invest. 105, 1353-1361. doi: 10.1172/JCI8862
Pubmed Abstract | Pubmed Full Text |Ref Full Text
Brekken, R. A., and Sage, E. H. (2001). SPARC, a matricellular protein: at the crossroads of cell-matrix communication. Matrix Biol. 19, 816-827. doi: 10.1016/S0945-053X(00)00133-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Carmeliet, P., and Jain, R. K. (2000). Angiogenese in kanker en andere ziekten. Nature 407, 249-257. doi: 10.1038/35025220
Pubmed Abstract | Pubmed Full Text |Ref Full Text
Chaudhury, A., Chander, P., and Howe, P. H. (2010). Heterogene nucleaire ribonucleoproteïnen (hnRNPs) in cellulaire processen: focus op de multifunctionele regulatorische rollen van hnRNP E1. RNA 16, 1449-1462. doi: 10.1261/rna.2254110
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Chaudhury, C., Mehnaz, S., Robinson, J. M., Hayton, W. L., Pearl, D. K., Roopenian, D. C., et al. (2003). De aan het major histocompatibility complex gerelateerde Fc-receptor voor IgG (FcRn) bindt albumine en verlengt de levensduur. J. Exp. Med. 197, 315-322. doi: 10.1084/jem.20021829
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Christensen, E. I., and Birn, H. (2002). Megalin en cubilin: multifunctionele endocytische receptoren. Nat. Rev. Mol. Cell Biol. 3, 256-266. doi: 10.1038/nrm778
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Commisso, C., Davidson, S. M., Soydaner-Azeloglu, R. G., Parker, S. J., Kamphorst, J. J., Hackett, S., et al. (2013). Macropinocytosis of protein is an amino acid supply route in Ras-transformed cells. Nature 497, 633-637. doi: 10.1038/nature12138
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Cui, S., Verroust, P. J., Moestrup, S. K., and Christensen, E. I. (1996). Megalin/gp330 medieert opname van albumine in renale proximale tubulus. Am. J. Physiol. 271, F900-F907. doi: 10.1038/nature12138
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Dennis, M. S., Zhang, M., Meng, Y. G., Kadkhodayan, M., Kirchhofer, D., Combs, D., et al. (2002). Albumin binding as a general strategy for improving the pharmacokinetics of proteins. J. Biol. Chem. 277, 35035-35043. doi: 10.1074/jbc.M205854200
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Desai, N., Trieu, V., Damascelli, B., and Soon-Shiong, P. (2009). SPARC expression correlates with tumor response to albumin-bound paclitaxel in head and neck cancer patients. Transl. Oncol. 2, 59-64. doi: 10.1593/tlo.09109
Pubmed Abstract | Pubmed Full Text |Ref Full Text
Desai, N. P., Trieu, V., Hwang, L. Y., Wu, R., Soon-Shiong, P., and Gradishar, W. J. (2008). Improved effectiveness of nanoparticle albumin-bound (nab) paclitaxel versus polysorbate-based docetaxel in multiple xenografts as a function of HER2 and SPARC status. Anticancer Drugs 19, 899-909. doi: 10.1097/CAD.0b013e32830f9046
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Evans, T. W. (2002). Review article: albumin as a drug-biological effects of albumin unrelated to oncotic pressure. Aliment. Pharmacol. Ther. 16(Suppl. 5), 6-11. doi: 10.1046/j.1365-2036.2002.00190.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Fritzsche, T., Schnolzer, M., Fiedler, S., Weigand, M., Wiessler, M., and Frei, E. (2004). Isolation and identification of heterogene nuclear ribonucleoproteins (hnRNP) from purified plasma membranes of human tumor cell lines as albumin-binding proteins. Biochem. Pharmacol. 67, 655-665. doi: 10.1016/j.bcp.2003.09.027
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Ghinea, N., Eskenasy, M., Simionescu, M., and Simionescu, N. (1989). Endothelial albumin binding proteins are membrane-associated components exposed on the cell surface. J. Biol. Chem. 264, 4755-4758.
Pubmed Abstract | Pubmed Full Text
Ghinea, N., Fixman, A., Alexandru, D., Popov, D., Hasu, M., Ghitescu, L., et al. (1988). Identification of albumin-binding proteins in capillary endothelial cells. J. Cell Biol. 107, 231-239.
Pubmed Abstract | Pubmed Full Text
Gradishar, W. J., Tjulandin, S., Davidson, N., Shaw, H., Desai, N., Bhar, P., et al. (2005). Fase III studie van nanodeeltjes-albumine-gebonden paclitaxel vergeleken met paclitaxel-gebaseerd op polyethylated castor oil bij vrouwen met borstkanker. J. Clin. Oncol. 23, 7794-7803. doi: 10.1200/JCO.2005.04.937
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Graham, J. M. (2002). Preparation of crude subcellular fractions by differential centrifugation. ScientificWorldJournal 2, 1638-1642. doi: 10.1100/tsw.2002.851
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Grasbeck, R. (2006). Imerslund-Grasbeck syndroom (selectieve vitamine B(12) malabsorptie met proteïnurie). Orphanet. J. Rare Dis. 1:17. doi: 10.1186/1750-1172-1-17
Pubmed Abstract | Pubmed Full Text |Ref Full Text
Greish, K. (2007). Enhanced permeability and retention of macromolecular drugs in solid tumors: a royal gate for targeted anticancer nanomedicines. J. Drug Target. 15, 457-464. doi: 10.1080/10611860701539584
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Han, N., Li, W., and Zhang, M. (2013). De functie van het RNA-bindende eiwit hnRNP in kankermetastase. J. Cancer Res. Ther. 9(Suppl.), S129-S134. doi: 10.4103/0973-1482.122506
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hawkins, J. W., and Dugaiczyk, A. (1982). The human serum albumin gene: structure of a unique locus. Gene 19, 55-58. doi: 10.1016/0378-1119(82)90188-3
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
He, X. M., and Carter, D. C. (1992). Atomic structure and chemistry of human serum albumin. Nature 358, 209-215. doi: 10.1038/358209a0
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Hradec, J. (1958). Metabolism of serum albumin in tumor-bearing rats. Br. J. Cancer 12, 290-304. doi: 10.1038/bjc.1958.35
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Iancu, C., Mocan, L., Bele, C., Orza, A. I., Tabaran, F. A., Catoi, C., et al. (2011). Verbeterde laserthermische ablatie voor de in vitro behandeling van leverkanker door specifieke toediening van multiwalled koolstofnanobuisjes gefunctionaliseerd met humaan serumalbumine. Int. J. Nanomed. 6, 129-141. doi: 10.2147/IJN.S15841
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Iwamoto, T. (2013). Clinical application of drug delivery systems in cancer chemotherapy: review of the efficacy and side effects of approved drugs. Biol. Pharm. Bull. 36, 715-718. doi: 10.1248/bpb.b12-01102
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Jain, R. K. (2001). Normalizing tumor vasculature with anti-angiogenic therapy: a new paradigm for combination therapy. Nat. Med. 7, 987-989. doi: 10.1038/nm0901-987
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Kragh-Hansen, U. (1981). Moleculaire aspecten van ligand binding aan serum albumine. Pharmacol. Rev. 33, 17-53. doi: 10.1038/nm0901-987
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Kratz, F. (2008). Albumine als geneesmiddeldrager: ontwerp van prodrugs, geneesmiddelconjugaten en nanodeeltjes. J. Control. Release 132, 171-183. doi: 10.1016/j.jconrel.2008.05.010
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Kratz, F. (2010). Albumine, een veelzijdige drager in de oncologie. Int. J. Clin. Pharmacol. Ther. 48, 453-455. doi: 10.5414/CPP48453
Pubmed Abstract | Pubmed Full Text |Ref Full Text
Kudlowitz, D., and Muggia, F. (2014). Nanoparticle albumin-bound paclitaxel (nab-paclitaxel): extending its indications. Expert Opin. Drug Saf. 13, 681-685. doi: 10.1517/14740338.2014.910193
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Lancon, A., Delmas, D., Osman, H., Thenot, J. P., Jannin, B., and Latruffe, N. (2004). Human hepatic cell uptake of resveratrol: involvement of both passive diffusion and carrier-mediated process. Biochem. Biophys. Res. Comm. 316, 1132-1137. doi: 10.1016/j.bbrc.2004.02.164
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Lovejoy, D. B., Jansson, P. J., Brunk, U. T., Wong, J., Ponka, P., and Richardson, D. R. (2011). Antitumor activiteit van metaal-chelerende verbinding Dp44mT wordt gemedieerd door vorming van een redox-actief kopercomplex dat accumuleert in lysosomen. Cancer Res. 71, 5871-5880. doi: 10.1158/0008-5472.CAN-11-1218
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Maeda, H., Wu, J., Sawa, T., Matsumura, Y., and Hori, K. (2000). Tumor vascular permeability and the EPR effect in macromolecular therapeutics: a review. J. Control. Release 65, 271-284. doi: 10.1016/S0168-3659(99)00248-5
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Matsumura, Y., and Maeda, H. (1986). A new concept for macromolecular therapeutics in cancer chemotherapy: mechanism of tumoritropic accumulation of proteins and the antitumor agent smancs. Cancer Res. 46, 6387-6392.
Pubmed Abstract | Pubmed Full Text
Mendlovic, F., and Conconi, M. (2010). Calreticulin: een eiwit met vele facetten. Nature Educ. 4, 1.
Merlot, A. M., Kalinowski, D. S., and Richardson, D. R. (2013a). Novel chelators for cancer treatment: where are we now? Antioxid. Redox Signal. 18, 973-1006. doi: 10.1089/ars.2012.4540
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Merlot, A. M., Pantarat, N., Menezes, S. V., Sahni, S., Richardson, D. R., and Kalinowski, D. S. (2013b). Cellulaire opname van het antitumor middel Dp44mT vindt plaats via een carrier/receptor-gemedieerd mechanisme. Mol. Pharmacol. 84, 911-924. doi: 10.1124/mol.113.088393
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Merlot, A. M., and Richardson, D. R. (2014). Receptorherkenning en lysosomale targeting om de cytotoxiciteit te verbeteren van nieuwe antikankermiddelen die ijzer en koper binden. Vitam. Miner. 3:e125. doi: 10.4172/vms.1000e125
CrossRef Full Text
Milici, A. J., Watrous, N. E., Stukenbrok, H., and Palade, G. E. (1987). Transcytosis of albumin in capillary endothelium. J. Cell Biol. 105, 2603-2612.
Pubmed Abstract | Pubmed Full Text
Moestrup, S. K., Kozyraki, R., Kristiansen, M., Kaysen, J. H., Rasmussen, H. H., Brault, D., et al. (1998). The intrinsic factor-vitamine B12 receptor and target of teratogenic antibodies is a megalin-binding peripheral membrane protein with homology to developmental proteins. J. Biol. Chem. 273, 5235-5242. doi: 10.1074/jbc.273.9.5235
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Neesse, A., Frese, K. K., Chan, D. S., Bapiro, T. E., Howat, W. J., Richards, F. M., et al. (2014). SPARC onafhankelijke medicijnafgifte en antitumor effecten van nab-paclitaxel in genetisch gemanipuleerde muizen. Gut 63, 974-983. doi: 10.1136/gutjnl-2013-305559
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Ober, R. J., Martinez, C., Lai, X., Zhou, J., and Ward, E. S. (2004). Exocytosis of IgG as mediated by the receptor, FcRn: an analysis at the single-molecule level. Proc. Natl. Acad. Sci. U.S.A. 101, 11076-11081. doi: 10.1073/pnas.0402970101
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Peters, T. (1996). Alles over albumine: Biochemistry, Genetics and Medical Applications. San Diego, CA: Academic Press Limited.
Peterson, H. I., and Appelgren, K. L. (1973). Experimental studies on the uptake and rentention of labelled proteins in a rat tumor. Eur. J. Cancer 9, 543-547. doi: 10.1016/0014-2964(73)90142-4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Podhajcer, O. L., Benedetti, L. G., Girotti, M. R., Prada, F., Salvatierra, E., and Llera, A. S. (2008). De rol van het matricellulaire eiwit SPARC in de dynamische interactie tussen de tumor en de gastheer. Cancer Metastasis Rev. 27, 691-705. doi: 10.1007/s10555-008-9146-7
CrossRef Full Text
Porter, P.L., Sage, E.H., Lane, T.F., Funk, S.E., and Gown, A.M. (1995). Distributie van SPARC in normaal en neoplastisch menselijk weefsel. J. Histochem. Cytochem. 43, 791-800. doi: 10.1177/43.8.7622842
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Roopenian, D. C., and Akilesh, S. (2007). FcRn: de neonatale Fc-receptor wordt volwassen. Nat. Rev. Immunol. 7, 715-725. doi: 10.1038/nri2155
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Sarav, M., Wang, Y., Hack, B. K., Chang, A., Jensen, M., Bao, L., et al. (2009). Renale FcRn recupereert albumine maar vergemakkelijkt eliminatie van IgG. J. Am. Soc. Nephrol. 20, 1941-1952. doi: 10.1681/ASN.2008090976
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Schilling, U., Friedrich, E. A., Sinn, H., Schrenk, H. H., Clorius, J. H., and Maier-Borst, W. (1992). Design of compounds having enhanced tumor uptake, using serum albumin as a carrier-Part II. In vivo studies. Int. J. Rad. Appl. Instrum. B 19, 685-695. doi: 10.1016/0883-2897(92)90103-6
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Schnitzer, J. E. (1992). gp60 is an albumin-binding glycoprotein expressed by continuous endothelium involved in albumin transcytosis. Am. J. Physiol. 262, H246-H254. doi: 10.1016/1050-1738(93)90012-U
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Schnitzer, J. E. (1993). Update on the cellular and molecular basis of capillary permeability. Trends Cardiovasc. Med. 3, 124-130. doi: 10.1016/1050-1738(93)90012-U
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Schnitzer, J. E., Allard, J., and Oh, P. (1995). NEM inhibits transcytosis, endocytosis, and capillary permeability: implication of caveolae fusion in endothelia. Am. J. Physiol. 268, H48-H55.
Pubmed Abstract | Pubmed Full Text
Schnitzer, J. E., and Bravo, J. (1993). High affinity binding, endocytosis, and degradation of conformationally modified albumins. Potential role of gp30 and gp18 as novel scavenger receptors. J. Biol. Chem. 268, 7562-7570.
Pubmed Abstract | Pubmed Full Text
Schnitzer, J. E., Carley, W. W., and Palade, G. E. (1988). Albumine interageert specifiek met een 60-kDa microvasculair endotheliaal glycoproteïne. Proc. Natl. Acad. Sci. U.S.A. 85, 6773-6777. doi: 10.1073/pnas.85.18.6773
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Schnitzer, J. E., and Oh, P. (1992). Antilichamen tegen SPARC remmen de binding van albumine aan SPARC, gp60, en microvasculair endotheel. Am. J. Physiol. 263, H1872-H1879.
Pubmed Abstract | Pubmed Full Text
Schnitzer, J. E., and Oh, P. (1994). Albondine-gemedieerde capillaire permeabiliteit voor albumine. Differential role of receptors in endothelial transcytosis and endocytosis of native and modified albumins. J. Biol. Chem. 269, 6072-6082.
Pubmed Abstract | Pubmed Full Text
Schnitzer, J. E., Sung, A., Horvat, R., and Bravo, J. (1992). Preferentiële interactie van albumine-bindende proteïnen, gp30 en gp18, met conformationeel gewijzigde albumines. Aanwezigheid in vele cellen en weefsels met een mogelijke rol in katabolisme. J. Biol. Chem. 267, 24544-24553.
Pubmed Abstract | Pubmed Full Text
Seetharam, B., Christensen, E. I., Moestrup, S. K., Hammond, T. G., and Verroust, P. J. (1997). Identification of rat yolk sac target protein of teratogenic antibodies, gp280, as intrinsic factor-cobalamin receptor. J. Clin. Invest. 99, 2317-2322. doi: 10.1172/JCI119411
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Sinn, H., Schrenk, H. H., Friedrich, E. A., Schilling, U., and Maier-Borst, W. (1990). Design of compounds having an enhanced tumor uptake, using serum albumin as a carrier. Deel I. Int. J. Rad. Appl. Instrum. B 17, 819-827. doi: 10.1016/0883-2897(92)90103-6
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Socinski, M. A., Bondarenko, I., Karaseva, N. A., Makhson, A. M., Vynnychenko, I., Okamoto, I., et al. (2012). Wekelijks nab-paclitaxel in combinatie met carboplatine versus oplosmiddelhoudende paclitaxel plus carboplatine als eerstelijnstherapie bij patiënten met gevorderde niet-kleincellige longkanker: eindresultaten van een fase III-studie. J. Clin. Oncol. 30, 2055-2062. doi: 10.1200/JCO.2011.39.5848
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Stehle, G., Sinn, H., Wunder, A., Schrenk, H. H., Stewart, J. C., Hartung, G., et al. (1997). Plasma proteïne (albumine) katabolisme door de tumor zelf-implicaties voor tumormetabolisme en het ontstaan van cachexie. Crit. Rev. Oncol. Hematol. 26, 77-100. doi: 10.1016/S1040-8428(97)00015-2
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Sudlow, G., Birkett, D. J., and Wade, D. N. (1976). Further characterization of specific drug binding sites on human serum albumin. Mol. Pharmacol. 12, 1052-1061.
Pubmed Abstract | Pubmed Full Text
Sugio, S., Kashima, A., Mochizuki, S., Noda, M., and Kobayashi, K. (1999). Kristalstructuur van menselijk serumalbumine bij 2.5 A resolutie. Protein Eng. 12, 439-446. doi: 10.1093/protein/12.6.439
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Tiruppathi, C., Finnegan, A., and Malik, A. B. (1996). Isolation and characterization of a cell surface albumin-binding protein from vascular endothelial cells. Proc. Natl. Acad. Sci. U.S.A. 93, 250-254. doi: 10.1073/pnas.93.1.250
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Tiruppathi, C., Song, W., Bergenfeldt, M., Sass, P., and Malik, A. B. (1997). Gp60 activation mediates albumin transcytosis in endothelial cells by tyrosine kinase-dependent pathway. J. Biol. Chem. 272, 25968-25975. doi: 10.1074/jbc.272.41.25968
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
van der Veldt, A. A., Luurtsema, G., Lubberink, M., Lammertsma, A. A., and Hendrikse, N. H. (2008). Individualized treatment planning in oncology: role of PET and radiolabelled anticancer drugs in predicting tumor resistance. Curr. Pharm. Des. 14, 2914-2931. doi: 10.2174/138161208786404344
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Vuignier, K., Schappler, J., Veuthey, J. L., Carrupt, P. A., and Martel, S. (2010). Drug-protein binding: een kritisch overzicht van analytische tools. Anal. Bioanal. Chem. 398, 53-66. doi: 10.1007/s00216-010-3737-1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Wang, J., Ueno, H., Masuko, T., and Hashimoto, Y. (1994). Binding van serumalbumine op tumorcellen en karakterisering van het albumin bindend eiwit. J. Biochem. 115, 898-903.
Pubmed Abstract | Pubmed Full Text
Wunder, A., Stehle, G., Sinn, H., Schrenk, H., Hoffbiederbeck, D., Bader, F., et al. (1997). Verbeterde albumine-opname door rattentumoren. Int. J. Oncol. 11, 497-507. doi: 10.3892/ijo.11.3.497
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Yamagishi, T., Sahni, S., Sharp, D. M., Arvind, A., Jansson, P. J., and Richardson, D. R. (2013). P-glycoproteïne medieert geneesmiddelenresistentie via een nieuw mechanisme waarbij lysosomale sequestratie betrokken is. J. Biol. Chem. 288, 31761-31771. doi: 10.1074/jbc.M113.514091
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text
Zhai, X. Y., Nielsen, R., Birn, H., Drumm, K., Mildenberger, S., Freudinger, R., et al. (2000). Cubilin- en megalin-gemedieerde opname van albumine in gekweekte proximale tubuulcellen van de nier van opossum. Kidney Int. 58, 1523-1533. doi: 10.1046/j.1523-1755.2000.00314.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text