Introduction
育種家や研究者は、子孫の生態的分岐につながる潜在的なドライバーとして、分岐した遺伝子型間の交雑や内殖に関心を持っています(Rieseberg et al.)
F2以降の世代で低下した雑種生存率および/または繁殖力の分離は雑種破壊(HB)と呼ばれ、劣性対立遺伝子は必ず関連する。 この生殖障害は植物でも動物でも古くから観察されている(Dobzhansky, 1970; Grant, 1971)。 多くの遺伝学的解析により、この障壁は対立遺伝子が分化した遺伝子座間および/または遺伝子座同士の相補効果によって達成されることが明らかになっており、一般にBateson-Dobzhansky-Muller (BDM) 非適合と呼ばれている(Rieseberg and Willis, 2007)。 HBは必然的に雑種生殖不能(弱体化、壊死、クロロシスなど)や雑種不妊(雄、雌、両方の配偶子)など、接合後の生殖障害を内在するため、F1子孫におけるHBと不妊の区別がつかない研究者もいるようである。 それでも、HBはその根底にある遺伝的基盤(すなわち、劣性対立遺伝子の関与)を示唆しているため、生殖障害の分類として便利だと考えている」
モデル植物種シロイヌナズナを用いた最近の研究により、HBについてその分子機構に関する理解が深まっている(Vaid and Laitinen,2019)。 逆に,イネ(Oryza種)はモデル作物であるが,この種ではHBの遺伝的基盤に関する現在の理解はまだ限られており,それはおそらく,親となる近交系と比較して高い穀物生産が期待できるF1雑種ではHBが生殖障壁とならず,F2以降の世代で分離する劣った植物は育種集団から表現型に基づいて容易に選択できるためと思われる。
ここでは、シロイヌナズナの研究成果を参考にしながら、イネの交配におけるHBについて概説し、イネの育種においてHBがどのように扱われているかを論じる。
イネにおける雑種崩壊の遺伝的基盤
栽培イネであるジャポニカとインディカは、その祖先とされるルフィポゴンや野生種と種複合体を形成すると考えられているが、それらの交配ではHBなど複数の生殖障害が認められる(岡、1988; Vaughan et al, 2003)。
私の知る限り、岡博士はジャポニカとインディカの亜種間交配におけるHBの遺伝的基盤を初めて記述し、それは2つの相補的遺伝子によって説明された(Oka, 1957)。
イネの交配では、亜種間(O. sativa ssp. japonica × ssp. indica)および種間(O. sativa × O. nivara and O. sativa × O. glumaepatula)交配でHBがしばしば記載されている(岡、1957;佐藤および森島、1988;Wuら、.1987)。 1995; Li et al., 1997; Fukuoka et al., 1998, 2005; Kubo and Yoshimura, 2002, 2005; Matsubara et al., 2007a,b, 2015; Yamamoto et al., 2007; Ichitani et al., 2012; for intersubspecific crosses; Sobrizal et al., 2001; Miura et al., 2008) (Table 1). これらのHB例の多くは2座BDM不適合と劣性対立遺伝子によるもので、両座で劣性対立遺伝子をホモ接合で持つF2子孫の1/16がHB表現型を示し、一方、1座のみでヘテロ接合である子孫の4/16がHB表現型を示し、交配の組み合わせにより異なる(表1および図1A)。 戻し交配雑種では、1つの遺伝子座がすでに再帰親からの対立遺伝子で固定されていた。したがって、BCnF2子孫の1/4がHB表現型を示した(表1および図1B)

Table 1. イネの交配で報告されたハイブリッドブレークダウンの遺伝的基盤.
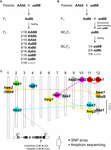
Figure 1.ハイブリッドブレークダウンの遺伝的基盤.
イネ雑種におけるHB表現型は交配組み合わせにより変化する。 例えば、hbd2/hbd2 hbd3/hbd3遺伝子型は弱さを示すが、明らかな種子不稔性はない。 しかし、hbd4/hbd4 hbd5/hbd5遺伝子型は、弱勢と種子不稔の両方を示す(Matsubara et al, 2007a, 2015; Yamamoto et al, 2007; Table 1)。 また、アソミノリ×IR24の交配という同じ交配組み合わせにもかかわらず、弱性と種子不稔の両方を示すHB(hwe1/hwe1 hwe2/hwe2 genotype)や種子不稔のみを示すHB(hsa1/hsa1 hsa2/hsa2 hsa3/hsa3 )が見られる複雑なケースもある(Kubo and Yoshimura, 2002, 2005; Table 1)。 これらのHBは独立した遺伝的基盤を持っているので、この交配ではF2子孫の少なくとも5/64がHB表現型を示すことになる。
現在までに、ジャポニカ米とインディカ米の交配において、DNAマーカーを用いていくつかのHBの原因遺伝子座が特定のゲノム領域にマッピングされている(Chr 7 and 10, Fukuoka et al, 1998; Chrs 1 and 12, Wu et al., 1995; Kubo and Yoshimura, 2002; Matsubara et al., 2015; Chrs 6 and 11, Fukuoka et al., 2005; Chrs 8, 9, and 12, Kubo and Yoshimura, 2005; Chrs 2 and 11, Matsubara et al. 2007a; Yamamoto et al. 2007, 2010; Chrs 11 and 12, Ichitani et al., 2012; Table 1 and Figure 1C). これらの研究の結果、イネのHBを支える遺伝子座は、ある交配では共有されているが、他の交配では異なっており、イネゲノムにおける多様化を示唆していることが明らかになった。 例えば、インシカ系品種に近縁なO. nivaraが持つHB対立遺伝子はインディカ系品種と同じ遺伝子座を共有している場合があるが、栽培品種に近縁なO. glumaepatulaが持つHB対立遺伝子は他の品種と共有していない(表1および図1C)。 なお、上記のHB例は、大きな影響を持つ一組の遺伝子によるものであるが、イネの育種において見過ごすことのできない、例えば、種子稔性がわずかに低下するなどの小さな影響を持つ一組の遺伝子による例もあり、まだ検出されていない。
Molecular Mechanisms Underlying Hybrid Breakdown
Autoimmune response: Bombliesら(2007)は、種内交配においてNB-LRR病害抵抗性遺伝子やR遺伝子がシロイヌナズナのHBと関連していることを初めて報告したが、この自己免疫反応は主にF1植物でネクロシスとして発現し、F2子孫のHBは温度によって条件付けられるようであった。 シロイヌナズナの交配種では、自己免疫反応によるHBを示すいくつかの証拠が報告されている(Alcázar et al., 2009, 2010)。 HBにNB-LRR遺伝子が関与していることから、複数のゲノム領域がシロイヌナズナのHBに関連し得ることが示唆され、実際、Chaeら(2014)が行った広範な調査はこの考えを支持している。
またイネの交配種では、自己免疫反応によるHBが、ジャポニカ×インディカ交配で報告されている(山本ら,2010). この場合、カゼインキナーゼIをコードし、インディカ品種が持つハイブリッドブレークダウン2(hbd2)遺伝子とジャポニカ品種が持つR遺伝子群が結合してF2植物にHBが発生したものである。 これらの原因対立遺伝子は部分劣性として振る舞う可能性が高いため、遺伝子産物が関与している可能性がある(Matsubara et al., 2007a; Yamamoto et al., 2007, 2010)。
重複遺伝子の相互抑制:この概念は雑種不適合性の遺伝的基盤として最初に理論的に提案された(WerthとWindham, 1991; LynchとForce, 2000)。 このモデルでは、一方の集団は一方の遺伝子座の機能を失い、他方の遺伝子座の機能を維持するが、他方の集団は逆の効果を経験する。 その結果、F2接合体の1/16は機能的な遺伝子を持たなくなる(Lynch and Force, 2000)。 Bikardら(2009)やVladら(2010)が報告したシロイヌナズナハイブリッドのHBは、この現象の良い例である。 このシナリオでは、エピジェネティックサイレンシングが関与するケースも報告されている(Agorio et al., 2017; Blevins et al., 2017)。
イネは全ゲノムとセグメント重複の両方を経験しているため(Wang et al., 2007; Guo et al., 2019)、ハイブリッド不適合の原因として相互サイレンシングが浸透しているかもしれない。 しかし、現在までのところ、イネにおいて重複遺伝子の相互サイレンシングによって引き起こされるHBの証拠はない。
自己免疫反応と相互サイレンシングとは無関係に、シロイヌナズナのハイブリッドでHBを引き起こすDEAD-box RNA helicase 18と別の遺伝子(有力候補としてMORPHEUS MOLECULE 1)の相互関係が明らかにされている(Plötner et al.、2017;Vaid et al.、2020)。 興味深いことに、HB表現型はF3世代とF4世代の間で減少し、遺伝子発現のエピジェネティックな制御を示唆している。
イネハイブリッドでは、3つのハイブリッドステリティ-a(hsa)遺伝子座のうち、Kuboら(2016)は最近、hsa1遺伝子座が2つの遺伝子からなり、これらの遺伝子はそれぞれDUF1618タンパク質とヌクレオチド結合タンパク質に若干類似性を有する未確認タンパク質とをコードすることを明らかにした。
従来のイネ育種におけるハイブリッド破壊
数十年にわたり、従来のイネ育種ではバルクポピュレーション法が広く採用されていた。 この方法では、交配後、F4またはF5集団は(単一種子降下法ではなく)人工選抜を行わずにバルクで自家受精させるが、初期世代集団は自然選抜と生存率選抜を行う(詳細はAllard, 1960; Ikehashi and Fujimaki, 1980)。
ドナーから品種への望ましい形質の導入や蓄積のために、イネ育種家はしばしば戻し交配や多重親交配を行い、その後にバルクポピュレーション法を用いてきた。 経験的に、これらの交配法は、雑種子孫におけるHBに関連する対立遺伝子の破壊的組み合わせの分離をしばしば減少させるため、生殖障壁による選択候補の損失を緩和できることが長い間知られてきた。 しかし、これらの交配法による生殖障壁の緩和は、必然的に両親のHB遺伝子型に依存する。
ゲノミクスアサルト育種におけるハイブリッド破壊
基準作物ゲノム配列の公開と次世代シーケンサー技術の開発は、作物の分子育種の進展を加速した(Kole et al, 2015). こうした中,作物育種ではゲノムワイドDNAマーカーの遺伝子型に基づくゲノム選抜などのゲノムアシスト育種が検討されている(Spindel and Iwata, 2018)
自家受粉作物のゲノムアシスト育種では,組み換え近交系などの先進世代集団を参照集団として使用し,そこからゲノムワイドな遺伝子型・表現型データを得ることが多い。 その後、マーカー遺伝子型のみに基づく選抜が、初期世代の子孫(例えば、F2)において行われる。 この選抜法では、交配後の初期世代で弱い子孫や不稔の子孫は排除されているはずなので、参照集団は通常、HBのゲノム領域の情報を提供しないことに注意する必要がある。 したがって、このような選抜方式を採用した場合、いくつかの重要な選抜候補を捨てなければならない。
討論
このテーマに関する研究の数は限られているが、イネの交配におけるHBの基礎となる遺伝子座は共有というより多様化しているようである(表1および図1)。 以上のように、交配親間のHB関連対立遺伝子の分布に関する情報は、イネの育種、特にゲノム支援育種の効果を最大限に発揮するための必須条件となるはずである。 したがって、イネ交配におけるHBの追加検出と、イネゲノムにおける責任遺伝子のマッピングが必要である。 シロイヌナズナの交配種では、R遺伝子とその相互作用遺伝子に関するそのようなデータが広く調査されている(Alcázar et al.) 山本ら(2010)によるイネハイブリッドでのhbd2遺伝子の報告は、SNPアレイやアンプリコンシーケンスを用いて行われた広範な調査の良い例である(図1C)。 hbd3遺伝子やhsa1遺伝子も、この種の調査の候補である。 最終的には、親系統のゲノム情報に基づいてHBを予測できるような基準を開発することが、ゲノムアシスト育種の重要なツールとなるであろう。 また、原因遺伝子が単一遺伝子として特定されない場合でも、栽培イネの連鎖不平衡は100-200kbに及ぶと推定されていることから、一塩基多型などの密接に結びついたマーカーは、HBに関連するハプロタイプの分類に有効であろう(野生種(O.M.)などでは、連鎖不平衡は100-200kbに及ぶと推定されている)。 rufipogon)は数十kbに及ぶ可能性がある(Mather et al., 2007; McNally et al., 2009; Huang et al., 2010)。 この情報は、より効率的で効果的な交配組合せの設計を可能にする。
バルク分離解析に続いて次世代シーケンサーを用いることで、従来の連鎖地図作成(量的形質遺伝子座解析など)と同様に、HB分離体が各交配集団における通常生育分離体と区別できる特徴(植物高、ティラー数、豊饒などの形質について)を示すことが多いため、HB関連座位のマッピングに役立つと期待される。 また、デジタルカメラやドローン搭載による画像データの取得も、実験室や水田での表現型決定に重要な役割を果たすと思われる。 これらの取り組みは、イネ育種家だけでなく、進化生物学者にも貴重な情報を提供するだろう。
さらに、HB関連対立遺伝子が特定された場合、イオンビームやゲノム編集による遺伝子破壊が、イネのF1雑種不稔に代表されるようにHB克服に役立つかもしれない(小出ら、2018;Xie et al, 2019)。
Data Availability Statement
本研究で提示されたすべてのデータセットは、論文/補足資料に含まれている。
Author Contributions
著者は、この作品の唯一の貢献者であることを認め、出版を承認している。
Conflict of Interest
著者は、潜在的な利益相反と解釈される商業的または金銭的関係がない状態で研究が行われたことを宣言する。
Acknowledgements
私はこの論文を、ダルシャン・ブラール博士の思い出に捧げたいと考えている。 彼は、野生種が持っていた有用な遺伝子を、生殖の壁を超えて品種に精力的に導入した著名な稲育種家である。
Allard, R. W. (1960). 植物育種の原理(Principles of Plant Breeding). ニューヨーク、NY: John Wily and Sons, Inc.
Google Scholar
Dobzhansky, T. (1970). 進化過程の遺伝学. ニューヨーク、NY: コロンビア大学出版会。
Google Scholar
Grant, V. (1971)。 植物の種分化. ニューヨーク、NY:
Google Scholar
Ikehashi, H., and Fujimaki, H. (1980). “Modified bulk population method for rice breeding,” in Proceedings of the International Rice Research Conference on Innovative approaches to rice breeding, (Los Baños, PA: International Rice Research Institute), 163-182.
Google Scholar
Oka, H.-I. (1988). 栽培米の起源. 東京. JSSP.
Google Scholar
.