評価|生物心理学|比較|認知|発達|言語|個人差|性格|哲学|社会|
方法|統計|臨床|教育|産業|専門項目|世界心理|
生物学的。行動遺伝学-進化心理学-神経解剖学-神経化学-神経内分泌学-神経科学-精神神経免疫学-生理心理学-精神薬理学(索引、概要)
| Brain: 視床下部 | ||
|---|---|---|
 |
||
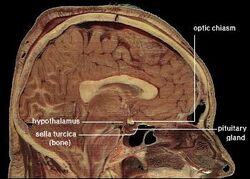
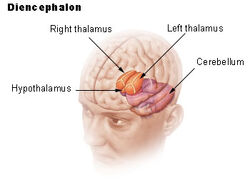
| ヒト視床下部の位置 | ||
 |
||
| Dienchephalon | ||
| Latin | hypothalamus | |
| グレーの | subject #189 812 | |
| 部分的な の | ||
| 成分 | ||
| 血管 | ||
| 脳情報/UW | ヒア(-)358 | |
| MeSH | A08.186.211.730.385.357 | |
視床下部は下垂体(hypophysis)を介して神経系を内分泌系にリンクしています。 視床下部(ギリシャ語のὑποθαλαμος=視床の下)は、視床の下、脳幹のすぐ上に位置しています。 この腺は、間脳の腹部の大部分を占めている。 すべての哺乳類の脳に存在する。 視床下部は、自律神経系の代謝過程とその他の活動を制御しています。 視床下部は、体温、飢餓、渇き、疲労、怒り、概日リズムを制御している。
入力
視床下部はヒトの脳の中で非常に複雑な領域で、視床下部の中の小さな核でさえ、多くの異なる機能に関与している。 例えば室傍核には、下垂体後葉に投射するオキシトシンやバソプレシン(抗利尿ホルモンともいう)ニューロンがあるが、ACTHやTSH分泌(下垂体前葉に投射)、胃の反射、母性行動、血圧、摂食、免疫反応、体温などを調節するニューロンもある。
視床下部は、多くのホルモンおよび行動の概日リズム、神経内分泌出力の複雑なパターン、複雑な恒常性維持機構、および多くの重要な行動を調整する。 そのため、視床下部は多くの異なるシグナルに反応する必要があり、その中には外部から発生するものと内部から発生するものがある。 そのため、視床下部は、脳幹網様体および自律神経ゾーン、大脳辺縁系(特に扁桃体、中隔、ブローカの対角帯、嗅球、大脳皮質)など、CNSの多くの部分と密接に関連している。
視床下部は、次のことに反応する:
- 光。
- フェロモンなどの嗅覚刺激
- 性腺ステロイドや副腎皮質ステロイドなどのステロイド
- 特に心臓や胃から生じる神経伝達情報
- 視床下部の反応:
- 光刺激(日長と光周期)
- 光刺激(フェロモンや副腎皮質ステロイドを含む。 生殖器<9774><3497>自律神経入力<9774><3497>血中刺激(レプチン、グレリン、アンジオテンシン、インスリン、下垂体ホルモン、サイトカイン、グルコースの血漿濃度、浸透圧など)<9774><3497>ストレス<9774><3497>体温上昇による微生物の侵入、体温調節器の上昇をリセットする。
嗅覚刺激
嗅覚刺激は多くの種で生殖や神経内分泌機能に重要である。 例えば、妊娠中のマウスが交尾後の重要な時期に「見知らぬ」オスの尿にさらされると、妊娠は失敗に終わります(ブルース効果)。 フェロモンによる合図は、多くの種で発情の同期化を助けます。女性の場合、月経の同期化もフェロモンによる合図から生じるかもしれませんが、人間におけるフェロモンの役割については疑問視されています。
血液による刺激
ペプチドホルモンは視床下部に重要な影響を与えるが、そのためには血液脳関門を通過する必要がある。 視床下部は、有効な血液脳関門を持たない特殊な脳領域に一部囲まれており、これらの部位の毛細血管内皮は、大きなタンパク質や他の分子でさえ自由に通過できるように柵状になっています。 このような部位には、神経下垂体や正中隆起といった神経分泌の場がある。 しかし、それ以外の部位は、脳が血液の成分を採取する場所である。 これらの部位のうち、角下器官とOVLT(末端薄層の血管器官)は、神経細胞が血液と髄液の両方に密接に接している、いわゆる周産期器官である。 これらの構造体は高密度に血管が張り巡らされており、飲水、バソプレシン放出、ナトリウム排泄、ナトリウム食欲を制御するオスモレセプターおよびナトリウムレセプターニューロンを含んでいる。 また、アンジオテンシン、心房性ナトリウム利尿因子、エンドセリンおよびリラキシンに対する受容体を持つニューロンも含まれており、これらはそれぞれ体液および電解質バランスの調節において重要である。 OVLT と SFO のニューロンは、視床上部の核と室傍核、および視床下部前部へ投射しています。 また、室傍ニューロンへの影響を介して、発熱とACTH分泌の両方を誘発するインターロイキンの作用部位である可能性もある。
視床下部の活動に影響を与えるすべてのペプチドが、どのようにして必要なアクセスを獲得しているかは明らかではない。 プロラクチンやレプチンの場合、血液からCSFへの脈絡叢での積極的な取り込みが証明されている。 下垂体ホルモンの中には、視床下部の分泌に負のフィードバック影響を及ぼすものがある;例えば、成長ホルモンは視床下部にフィードバックされるが、どのようにして脳に入るかは不明である。 また、プロラクチンおよびTSHの中枢作用に関する証拠もある。
ステロイド
視床下部には、性腺ステロイドおよびグルココルチコイド(ACTHに反応して放出される副腎のステロイドホルモン)に感応する神経細胞がある。 また、視床下部には、食欲に重要なグルコース感受性の特殊なニューロン(視床弧状核と視床下部下内側)もあります。 視床下部は脳幹から多くの入力を受けており、特に孤束路核、青斑核、腹外側髄質からの入力が重要である。 哺乳または膣頸部刺激に応じたオキシトシン分泌はこれらの経路のいくつかによって媒介される。頸動脈洞および大動脈弓の化学受容体および低圧心房容量受容体から生じる心臓血管刺激に応じたバソプレシン分泌は、他の経路によって媒介される。 ラットでは、膣の刺激によってもプロラクチンが分泌され、不妊交配後の擬似妊娠につながる。 ウサギでは、交尾により反射的に排卵が誘発される。 ヒツジでは、高濃度のエストロゲンの存在下で子宮頸管を刺激すると、処女の雌羊に母性行動を誘発することができます。 これらの作用はすべて視床下部が仲介し、情報は主に脳幹を中継する脊髄経路によって運ばれる。
心血管系の刺激は迷走神経によって伝えられるが、迷走神経はまた、例えば胃の膨張から生じる摂食を抑制する信号など、さまざまな内臓情報を伝える。 この情報もまた、脳幹のリレーを経由して視床下部に到達する。
核
視床下部の核には以下のものがある。
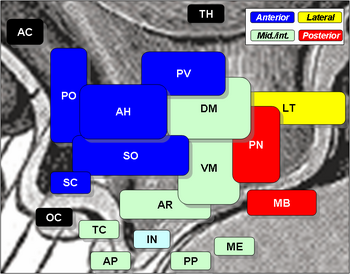
視床下部核
Region Area Nucleus 機能 前方 内側 内側視蓋前核 - 泌尿器系 膀胱収縮
- 心拍数減少
- 血圧低下
Supraoptic(視索上体 室傍核(SO) - バソプレシン放出
室傍核(PV) - オキシトシン放出
- バソプレシン放出
視床下部前核 (AH) - オキシトシン放出
- バソプレシン放出
バソプレシン放出 - 体温調節
- 喘ぎ
- 発汗
- チロトロピン抑制
視交叉上核(SC) - バソプレシン放出
- サーカディアンリズム
外側 外側視索前核 外側核(LT) - 渇きと空腹
視索上核(SO)の一部 - バソプレッシン 放出
視床下部背側核(DM) - GI 刺激
内側核(VM) - 飽和
- 神経内分泌コントロール
。
弧状核(AR) - 神経内分泌コントロール
外側 外側核(LT) - 口渇・空腹感
外側結節核 後 内側 乳頭核(乳頭体の一部)(MB) - 摂食 反射
Posterior nucleus (PN) - 血圧上昇
- 瞳孔の増加 dilation
- shivering
Lateral Lateral nucleus (LT) も参照されたい。 腹外側視索前核
出力
視床下部の出力は、神経投射と内分泌ホルモンの2つに分けられる。
神経投射
視床下部のほとんどの線維系は2方向に走っている(双方向性)。
- 視床下部の尾側領域への投射は内側前脳束、乳突片路、背外側縦束を経由している。
- 視床下部の吻側領域への投射は、乳頭視床路、線条体、終末線条体によって運ばれる。
内分泌ホルモン
視床下部は内分泌系に影響を与え、怒りや性的活動などの情動行動を支配している。 生成された視床下部ホルモンのほとんどは、視床下部門脈系を経由して下垂体に分配される。 視床下部は、血圧、心拍数および体温の調節を含む恒常性を維持する。
視床下部の主要なホルモンは以下の通りである。
名称 その他の名称 略号 所在地 機能 コルチコトロピン放出ホルモン コルチコトロピン放出因子 CRH,コルチコレブリン, CRF 室傍核の神経内分泌ニューロン バソプレシンと共に下垂体前葉を刺激してACTHを分泌する Dopamine Prolactin-inhibiting hormone DA, PIH 弧状核の神経内分泌ニューロン 下垂体前葉からのプロラクチン分泌を抑制する Gonadotropin Releasing hormone Luteinising-hormone Releasing hormone GnRH。 LHRH 視蓋内側前核および弧状核の神経内分泌ニューロン 下垂体前葉を刺激してLHおよびFSHを分泌する 成長ホルモン放出ホルモン 成長ホルモン放出因子 。 somatocrinin
GHRH、GHRF。 GRF 下垂体前葉神経内分泌ニューロン 刺激により成長ホルモンを分泌 メラトニン 視交叉上核 ソマトスタチン 成長ホルモン抑制するホルモン。 ソマトトロピン放出抑制因子 SS, GHIH, SRIF 脳室周囲核の神経内分泌ニューロン 下垂体前葉からの成長ホルモン分泌を抑制する チロトロピン放出ホルモン チロトロピン放出因子, チロリベリン、プロティレリン TRH, TRF 視床下部傍核および前部傍核の神経内分泌ニューロン stimulates anterior pituitary to seclee TSH 関連ページ:Thermos, TRF
TRH、Thermos STYLE、Thermos TSH、TRF、TRF
STYLE ヒポクレチン 食物摂取の制御
視床下部の腹内側核の極外側は、食物摂取の制御を担当する。 この部位を刺激すると食物の摂取量が増加する。 この部位の両側の病変は食物の摂取を完全に停止させる。 核の内側は外側を制御する作用がある。 腹内側核の内側部分の両側病変は動物の食欲亢進と肥満を引き起こす。
この調節に関連した異なる仮説がある:
- Lipostatic hypothesis – この仮説は、脂肪組織が脂肪量に比例した体液性シグナルを生成し、視床下部に作用して食物摂取量を減らしエネルギー出力を増加させるとするものである。 レプチンというホルモンが視床下部に作用して、食物摂取量を減少させ、エネルギー出力を増加させることが明らかになっている。
- 腸ペプチド仮説 – Grp、グルカゴン、CCKなどの消化管ホルモンが食物摂取を抑制すると主張するものである。 消化管に入った食物が引き金となってこれらのホルモンが放出され、脳に作用して満腹感をもたらす。 脳にはCCK-AとCCK-Bの両方の受容体が存在する。
- グルコスタティック仮説-腹内側核の満腹中枢の活性は、おそらく神経細胞のグルコース利用によって支配されている。 グルコース利用率が低く、その結果、ニューロン間の動静脈血中グルコース差が小さくなると、ニューロン間の活性が低下すると仮定されている。 このような状態では、摂食中枢の活動は抑制されず、空腹感を感じる。
- サーモスタット仮説-この仮説によると、体温が所定の設定値より下がると食欲が刺激され、逆に設定値より上がると食欲が抑制される。
Sexual dimorphism
視床下部のいくつかの核は性的に二型である、すなわち、構造と機能の両方で男性と女性の間に明確な違いがある。
いくつかの違いは総神経解剖でも明らかで、最も顕著なのは男性だけに存在する前頭葉内の性二型の核である。
これらの変化の重要性は、オスとメスの機能的な違いによって認識することができる。 例えば、成長ホルモンの分泌パターンは性差があり、これが多くの種で成体の雄が雌よりはるかに大きい理由の一つである。
卵巣ステロイドに対する反応
他の顕著な機能的二型は、成体の卵巣ステロイドに対する行動反応にある。 男性と女性では卵巣ステロイドに対する反応が異なるが、これは視床下部のエストロゲン感受性ニューロンの発現が性的に二型であること、すなわちエストロゲン受容体が異なるニューロンセットで発現することが一因である。
エストロゲンとプロゲステロンは特定のニューロンにおける遺伝子発現に影響を及ぼしたり、細胞膜電位の変化やキナーゼ活性化を誘導して、多様なノンゲノム細胞機能を引き起こすことができる。 エストロゲンやプロゲステロンは、それぞれの核内ホルモン受容体に結合し、細胞核に移動してホルモン応答要素(HRE)と呼ばれるDNA領域と相互作用したり、他の転写因子の結合部位にくくりつけられたりしている。 エストロゲン受容体(ER)は、遺伝子の近位プロモーター領域にエストロゲン応答要素(ERE)がないにもかかわらず、このように他の転写因子を転写活性化することが示されている。 ERとプロゲステロン受容体(PR)は一般に遺伝子活性化因子であり、ホルモン曝露後にmRNAとそれに続くタンパク質合成が増加する。
男性と女性の脳ではエストロゲン受容体の分布が異なり、この違いは新生児期のステロイド曝露の不可逆的な結果であるとされている。 エストロゲン受容体(およびプロゲステロン受容体)は、主に視床下部前部および中基部のニューロン、特に
- 視索前部(LHRHニューロンがある)
- 脳室周囲核(ソマトスタチンニューロンがある)
- 視床下部腹側(性的行動に重要である)に存在する。
ラット新生児期の性腺ステロイド
新生児期において、性腺ステロイドは神経内分泌系の視床下部の発達に影響を与える。 例えば、雌が正常な生殖周期を示すこと、雄と雌が成体で適切な生殖行動を示すことなどが決定される。
- 雌のラットに生後数日間(性ステロイドの影響が及ぶ「臨界期」)にテストステロンを1回注射すると、視床下部は不可逆的に男性化する。成体ラットは、エストロゲンに応答してLHサージ(雌の特徴)を生成できなくなるが、男性の性行動(性的受容力を持つ雌に乗りかかる)を示すことができるようになる。
- 対照的に、出生直後に去勢した雄ラットは雌化し、成体はエストロゲンに反応して雌の性行動(性的受容性、殿部形成)を示す。
霊長類のアンドロゲン
霊長類ではアンドロゲンの発達的影響はあまり明確ではなく、その結果はあまり完全ではないのです。 女の子の「おてんば娘」は胎児の脳へのアンドロゲンの影響を反映しているかもしれないが、最初の2-3年の間の飼育の性別は、ジェンダーアイデンティティの最も重要な決定要因であると多くの人が信じている。この段階では、エストロゲンまたはテストステロンのいずれかが女性または男性の脳のいずれかに永久的な影響を与え、異性愛と同性愛の両方に影響を与えるからだ。 脳内では、テストステロンはアロマ化して(エストラジオール)となり、発育に影響を与える主要な活性ホルモンとなる。 ヒトの精巣は、胎児期の約8週目から出生後5-6ヶ月まで高レベルのテストステロンを分泌し(多くの種で同様の周産期のテストステロン急増が観察される)、この過程が男性表現型の根底にあると思われる。
視床下部の発達に及ぼすその他の影響
視床下部の発達に重要な影響を及ぼすのは性ステロイドだけではない;特に、思春期前のストレスが、成人の視床下部の急性ストレスへの対応能力を決定する。 性腺ステロイド受容体とは異なり、グルココルチコイド受容体は脳全体に広く存在し、室傍核ではCRF合成と分泌の負のフィードバック制御を媒介しますが、その他の部位での役割はよく分かっていません。
加齢による視床下部への影響
雌マウスを用いた研究では、視索上核(SON)と室傍核(PVN)の両方が、正常な加齢によってIGF-1R免疫反応細胞のおよそ3分の1を失うことが示されています。 また、カロリー制限(CR)マウスでは、Old-Alマウスと比較して、IGF-1R免疫応答細胞数は同程度であるが、IGF-1R非免疫応答細胞数がより多く減少することがわかった。 その結果、Old-CRマウスは、正常な老化マウスと比較して、IGF-1に対する視床下部の感受性の増加を反映して、IGF-1R免疫反応性細胞の高いパーセンテージを示すことがわかった。
- HPA axis
- Hypothalamo hypophyseal system
- Hypothalamo pituitary adrenal system
- Hypothalamus lesions
- Medial Forebrain bundle
- 神経内分泌学
- 視索前野
も参照してください。
追加画像
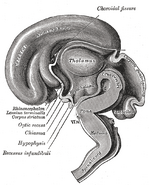 ヒト3ヶ月胚の脳の正中矢状断面図。
ヒト3ヶ月胚の脳の正中矢状断面図。このギャラリーに写真を追加する
- http://www.cancer.gov/Templates/db_alpha.aspx?CdrID=46359
- http://www.sci.uidaho.edu/med532/hypothal.htm
- 核の図 (psycheducation.jp)
- Diagram of Nuclei (universe-review.ca)
- Diagram of Nuclei (utdallas.edu)
- 表で特に指定がなければ、refは。 Guyton Eight Edition
- Walter F., PhD. ボロン メディカル・フィジオロジー A Cellular And Molecular Approaoch、Elsevier/Saunders。 840ページ
- http://thalamus.wustl.edu/course/hypoANS.html
- http://www.vivo.colostate.edu/hbooks/pathphys/endocrine/hypopit/overview.html
- Theologides A (1976). 食欲不振産生中間代謝産物。 Am J Clin Nutr 29 (5): 552-8.
- ジョン・マネー「39年後の幼年期・青年期の性同一性障害の概念」『性・夫婦療法研究』20 (1994): 163-77.
- Romeo, Russell D, Rudy Bellani, Ilia N. Karatsoreos, Nara Chhua, Mary Vernov, Cheryl D. Conrad and Bruce S. McEwen (2005). ストレス歴と思春期の発達が相互に作用して、視床下部-下垂体-副腎軸の可塑性を形成している。 Endocrinology 147 (4): 1664-1674.
- Saeed O,Yaghmaie F,Garan SA,Gouw AM,Voelker MA,Sternberg H,Timiras PS. (2007). インスリン様成長因子-1受容体免疫反応性細胞は、カロリー制限マウスの室傍視床下部に選択的に維持されている。 Int J Dev Neurosci 25 (1): 23-8.
- Yaghmaie F, Saeed O, Garan SA, Voelker MA, Gouw AM, Freitag W, Sternberg H, Timiras PS (2006). 視床下部上部のインスリン様成長因子-1受容体免疫反応細胞の年齢依存的な損失は、カロリー制限マウスで減少する。 Int J Dev Neurosci 24 (7): 431-6.
- F. Yaghmaie, O. Saeed, S.A. Garan, A.M. Gouw, P. Jafar, J. Kaur, S. Nijjar, P.S. Timiras, H. Sternberg, M.A. Voelker (2007). 加齢とカロリー制限による視床下部IGF-1感受性の変化を追跡する。 Experimental Gerontology 42 (1-2): 148-149.
参考文献 & 文献
論文
- Arees, E.A. and Mayer, B, (1967) Anatomical connections between medial and lateral regions of the hypothalamus concerned with food intake, Science 157:
- Grossman, S.P. (1960) Eating or drinking elicited by direct adrenergic or cholinergic stimulation of the hypothalamus, Science 132: 301-2.
- Hetherington, A.W. and Ranson, S.W. (1942) The relation of various hypothalamic lesions to adiposity in the rat, 76: 475-99.Grossman (1960) Adrenalic or Cholingic stimulation of the hypothalamus in direct adrenergic stimulation, Science 132: 301-5.
追加資料
論文
- Google Scholar
- BrainMaps at UCDavis 視床下部
- Endocrine system and hypothalamus
- High->BrainMaps at UCDavis霊長類の脳アトラス
- The Hypothalamus and Pituitary at endotexts.org
- Diagram of Nuclei (psycheducation.org)
- Diagram of Nuclei (universe-review.ca)
- Diagram of Nuclei (utdallas.org)
- Diagram of Nuclei (utdallas.org)edu)
v-d-e人間の脳:間脳(TA A14.1.08, GA 9.807)
視床下部 表面Pineal 体 – 羽根膜 – 羽根膜三角錐 – 羽根膜交連
前頭部 – 羽根膜核 – 下交連 器官
タラマス 表面ストライアイス 視床髄質-視床網状核-テニアタラミ
灰白質/
核対になる。 AN ・腹側(VA/VL, VP/VPM/VPL) ・外側(LD, LP, Pulvinar) ・後視床(MG, LG)
中線。 MD ・層内(Centromedian) ・正中核群 ・視床間接着
Mammillothalamic fasciculus ・Pallidothalamic tracts (Ansa lenticularis, Lenticular fasciculus, Thalamic fasciculus) ・PCML (Medial lemniscus, 棘突起視床路 – 側溝視床路 – 歯状視床路 – 音響放射 – 視神経放射 – 視床下筋膜 – 視床下部 – 前三叉神経節 路
髄層
視床下部 表面正中隆起/正中結節 – 乳頭体 – 下垂体
自律神経ゾーン前部(副交感神経・熱損失)-後部(交感神経・熱保存)
下垂体前部。 大細胞性/室傍性/上垂体性(オキシトシン/バソプレシン)
その他。 傍室性/弧状(ドーパミン/GHRH)-視索前部(GnRH)-視交叉上部(メラトニン)
外側(空腹感)-下内側(満腹感) – 背内側(怒り)
求心性(SN→内側前脳束)-求心性(乳頭状筋膜→AN, Stria terminalis → Amygdala, Dorsal longitudinal fasciculus → SC)
後部は間脳である。 ただし前方は腺
Subthalamus Subthalamic nucleus – Zona incerta
Ventricular system.がある。
第三脳室窪み。 (視床凹、下顎凹、上顎凹、松果体凹)
視床下部溝-第3脳室Tela chorioidea
角下器
開口部: 間脳/単脳
後交連
v-d-e人体構造、内分泌器官。 内分泌腺
視床下部/
下垂体軸Thyroid gland (Parafollicular cell.)の項を参照。 甲状腺上皮細胞、甲状腺峡、甲状腺葉、甲状腺ピラミッド)
副甲状腺(オキシフィル細胞。 主任細胞)Chromaffin cells
糸球体座 – 糸状体 – 網状体
Zuckerkandl – 大動脈体 – 頸動脈体
の臓器
精巣 – 卵巣 – 黄体
神経節— 。 正中隆起 – 下垂体茎 – 下垂体 – ニシン小体
Pars intermedia – Pars tuberalis – Pars distalis – アシドフィルス(Somatotropes, Lactotropes) – 好塩基球(Corticotropes、Gonadotropes。
松果体 松果体 – 小胞体
島嶼部 膵臓 アルファ細胞 – ベータ細胞 – デルタ細胞 – PP細胞 – イプシロン細胞
人の脳。 辺縁系
扁桃体 ・帯状回 ・姦淫回 ・海馬 ・視床下部 ・乳頭体 ・側坐核 ・前頭葉眼窩皮質 ・海馬傍回