Arviointi | Biopsykologia | Vertaileva | Kognitiivinen | Kehitys | Kieli | Yksilölliset erot | Yksilölliset erot | Persoonallisuus | Filosofia | Sosiaalinen |
Menetelmät | Tilastot | Kliininen | Kasvatus | Kasvatustiede | Työelämässä | Ammatilliset kohteet | Maailmanlaajuinen psykologian |
biologinen:Käyttäytymisgenetiikka – Evoluutiopsykologia – Neuroanatomia – Neurokemia – Neuroendokrinologia – Neurotieteet – Psykoneuroimmunologia – Fysiologinen psykologia – Psykofarmakologia(Hakemisto, pääpiirteittäin)
| Aivot: Hypotalamus | ||||
|---|---|---|---|---|
 |
||||
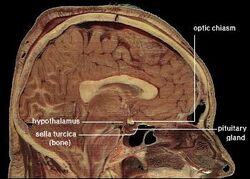
| Ihmisen hypotalamuksen sijainti | ||||
 |
||||
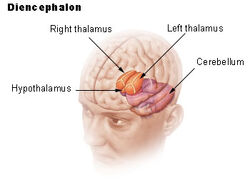
| Dienchephalon | ||||
| Latin | hypothalamus | |||
| Gray’s | subjektin #189 812 | |||
| Tietokanta #189 812 | ||||
| os. of | ||||
| Komponentit | ||||
| Arteria | ||||
| Veini | ||||
| Aivojen informaatio/UW | hier-358 | |||
| MeSH | A08.186.211.730.385.357 | |||
Hypotalamus yhdistää hermoston hormonitoimintaan aivolisäkkeen (hypofyysi) kautta. Hypotalamus, (kreikankielestä ὑποθαλαμος = talamuksen alla) sijaitsee talamuksen alapuolella, juuri aivorungon yläpuolella. Tämä rauhanen vie suurimman osan välikarsinan ventraalisesta alueesta. Sitä esiintyy kaikkien nisäkkäiden aivoissa. Ihmisellä se on suunnilleen mantelin kokoinen.
Hypotalamus säätelee tiettyjä aineenvaihduntaprosesseja ja muita autonomisen hermoston toimintoja. Se syntetisoi ja erittää neurohormoneja, joita kutsutaan usein hypotalamuksesta vapauttaviksi hormoneiksi, ja nämä puolestaan stimuloivat tai estävät aivolisäkehormonien eritystä.
Hypotalamus säätelee ruumiinlämpöä, nälkää, janoa, väsymystä, vihaa ja vuorokausirytmiä.
- Syötteet
- Hajuärsykkeet
- Verestä välittyvät ärsykkeet
- Steroidit
- Neuraaliset sisääntulot
- Ytimet
- Lähdöt
- Neuraaliset projektiot
- Endokriiniset hormonit
- Ruoan saannin kontrollointi
- Sukupuolidimorfismi
- Vastaukset munasarjojen steroideihin
- Gonaaliset steroidit rottien neonataalielämässä
- Androgeenit kädellisillä
- Muut vaikutukset hypotalamuksen kehitykseen
- Ikääntymisen vaikutukset hypotalamukseen
- Katso myös
- Lisäkuvat
- Viitteet & Kirjallisuusluettelo
- Artikkelit
- Lisämateriaali
- Artikkelit
Syötteet
Hypotalamus on hyvin monimutkainen alue ihmisen aivoissa, ja pienetkin ytimet hypotalamuksen sisällä osallistuvat moniin eri toimintoihin. Esimerkiksi paraventrikulaarinen ydin sisältää oksitosiinin ja vasopressiinin (jota kutsutaan myös antidiureettiseksi hormoniksi) neuroneja, jotka projisoituvat aivolisäkkeen takaosaan, mutta se sisältää myös neuroneja, jotka säätelevät ACTH:n ja TSH:n eritystä (jotka projisoituvat aivolisäkkeen etupuolelle), mahalaukun refleksejä, äitiyskäyttäytymistä, verenpainetta, ruokailua, immuunivasteet ja lämpötilaa.
Hypotalamus koordinoi monia hormonaalisia ja käyttäytymiseen liittyviä vuorokausirytmejä, neuroendokriinisten tuotosten monimutkaisia malleja, monimutkaisia homeostaattisia mekanismeja ja monia tärkeitä käyttäytymismalleja. Hypotalamuksen on siksi reagoitava moniin erilaisiin signaaleihin, joista osa syntyy ulkoisesti ja osa sisäisesti. Se on näin ollen tiiviissä yhteydessä moniin keskushermoston osiin, kuten aivorungon retikulaariseen muodostumaan ja autonomisiin vyöhykkeisiin, limbisiin etuaivoihin (erityisesti amygdalaan, septumiin, Brocan diagonaalikaistaleeseen ja hajulohkoihin sekä aivokuoreen).
Hypotalamus reagoi:
- valoon: Päivänpituus ja valojakso vuorokausi- ja vuodenaikarytmien säätelyyn
- Hajuärsykkeet, mukaan lukien feromonit
- Steroidit, mukaan lukien sukurauhassteroidit ja kortikosteroidit
- Neuronaalisesti välittyvä informaatio, joka syntyy erityisesti sydämestä, vatsasta, ja sukuelimistä
- Autonomiset tulot
- Verestä peräisin olevat ärsykkeet, mukaan lukien leptiini, greliini, angiotensiini, insuliini, aivolisäkkeen hormonit, sytokiinit, plasman glukoosipitoisuudet ja osmolaarisuus jne
- Sstressi
- Mikro-organismien tunkeutuminen elimistön lämpötilaa kohottamalla, jolloin elimistön lämpötermostaatti nollautuu ylöspäin.
Hajuärsykkeet
Hajuärsykkeet ovat tärkeitä lisääntymiselle ja neuroendokriiniselle toiminnalle monilla lajeilla. Jos esimerkiksi tiineenä oleva hiiri altistetaan ”vieraan” uroksen virtsalle kriittisenä aikana yhdynnän jälkeen, raskaus epäonnistuu (Brucen vaikutus). Naarashiiri muodostaa yhdynnän aikana tarkan ”hajumuistin” kumppanistaan, joka säilyy useita päiviä.Feromonivihjeet auttavat monilla lajeilla tiineyden synkronoinnissa; naisilla myös synkronoidut kuukautiset voivat johtua feromonivihjeistä, vaikka jotkut epäilevätkin feromonien merkitystä ihmisillä.
Verestä välittyvät ärsykkeet
Peptidihormoneilla on merkittäviä vaikutuksia hypotalamukseen, ja sitä varten niiden on ohitettava veri-aivoeste. Hypotalamus rajoittuu osittain erikoistuneisiin aivoalueisiin, joilta puuttuu tehokas veri-aivoeste; näissä kohdissa kapillaarien endoteeli on aukkoinen, jotta suuretkin proteiinit ja muut molekyylit pääsevät vapaasti läpi. Jotkin näistä alueista ovat neurosekretion paikkoja – neurohypofyysi ja median eminence. Toiset ovat kuitenkin kohtia, joissa aivot ottavat näytteitä veren koostumuksesta. Kaksi näistä paikoista, subfornikaalinen elin ja OVLT (organum vasculosum of the lamina terminalis) ovat niin sanottuja circumventrikulaarisia elimiä, joissa neuronit ovat läheisessä kosketuksessa sekä veren että CSF:n kanssa. Nämä rakenteet ovat tiheästi vaskularisoituneita, ja niissä on osmoreettisia ja natriumia vastaanottavaisia neuroneja, jotka säätelevät juomista, vasopressiinin vapautumista, natriumin erittymistä ja natriumin ruokahalua. Ne sisältävät myös neuroneja, joilla on reseptorit angiotensiinille, eteisnatriureettiselle tekijälle, endoteliinille ja relaksiinille, joista jokainen on tärkeä neste- ja elektrolyyttitasapainon säätelyssä. OVLT:n ja SFO:n neuronit projisoituvat supraoptiseen ytimeen ja paraventrikulaariseen ytimeen sekä preoptisiin hypotalamuksen alueisiin. Circumventrikulaariset elimet voivat olla myös interleukiinien vaikutuskohteena, joka saa aikaan sekä kuumeen että ACTH:n erityksen vaikuttamalla paraventrikulaarisiin neuroneihin.
Ei ole selvää, miten kaikki hypotalamuksen aktiivisuuteen vaikuttavat peptidit saavat tarvittavan pääsyn. Prolaktiinin ja leptiinin tapauksessa on näyttöä aktiivisesta imeytymisestä suonikalvopleksuksessa verestä CSF:ään. Joillakin aivolisäkkeen hormoneilla on negatiivinen takaisinkytkentä hypotalamuksen eritykseen; esimerkiksi kasvuhormoni vaikuttaa takaisin hypotalamukseen, mutta ei ole selvää, miten se pääsee aivoihin. Myös prolaktiinin ja TSH:n sentraalisesta vaikutuksesta on näyttöä.
Steroidit
Hypotalamus sisältää hermosoluja, jotka ovat herkkiä sukurauhassteroideille ja glukokortikoideille – (lisämunuaisen steroidihormonit, joita vapautuu vastauksena ACTH:lle). Se sisältää myös erikoistuneita glukoosille herkkiä neuroneja (kaarevassa ytimessä ja ventromediaalisessa hypotalamuksessa), jotka ovat tärkeitä ruokahalun kannalta. Preoptisella alueella on lämpöherkkiä neuroneja; nämä ovat tärkeitä TRH:n eritykselle.
Neuraaliset sisääntulot
Hypotalamus saa monia sisääntuloja aivorungosta; erityisesti yksinäisen radan ytimestä, locus coeruleuksesta ja ventrolateraalisesta medullasta. Oksitosiinin eritystä vasteena imemiseen tai vagino-cervikaaliseen stimulaatioon välittävät jotkin näistä reiteistä; vasopressiinin eritystä vasteena kardiovaskulaarisiin ärsykkeisiin, jotka johtuvat sinus carotiksen ja aortan kaaren kemoreseptoreista ja matalapaineisista eteisen tilavuusreseptoreista, välittävät toiset. Rotilla emättimen stimulaatio aiheuttaa myös prolaktiinin eritystä, ja tämä johtaa hedelmättömän parittelun jälkeiseen näennäisraskauteen. Kaniinilla yhdyntä saa aikaan refleksinomaisen ovulaation. Lampaalla kohdunkaulan stimulaatio korkeiden estrogeenipitoisuuksien läsnä ollessa voi saada aikaan neitsytuoreen uuhen äidin käyttäytymisen. Kaikki nämä vaikutukset välittyvät hypotalamuksesta, ja tiedot kulkevat pääasiassa aivorunkoon välittyvien selkäydinreittien kautta. Nännien stimulointi stimuloi oksitosiinin ja prolaktiinin vapautumista ja hillitsee LH:n ja FSH:n vapautumista.
Kardiovaskulaariset ärsykkeet välittyvät vagushermon kautta, mutta vagus välittää myös monenlaista viskeraalista informaatiota, mukaan lukien esimerkiksi mahalaukun pullistumisesta johtuvat signaalit ruokinnan hillitsemiseksi. Nämäkin tiedot saapuvat aivorungossa olevien releiden kautta hypotalamukseen.
Ytimet
Hypotalamuksen ytimiin kuuluvat seuraavat:
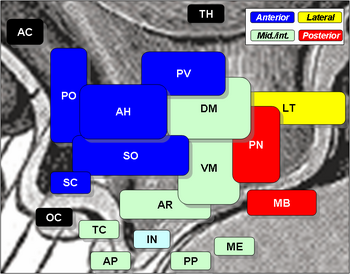
Hypotalamuksen ytimet
| Regio | Alue | Ydin | Tehtävä |
| Anteriorinen | Mediaalinen | Mediaalinen preoptinen ydin |
|
| Supraoptinen nucleus (SO) |
|
||
| Paraventrikulaarinen ydin (PV) |
|
||
| Anteriorinen hypotalamuksen ydin (AH) |
|
||
| Suprachiasmaattinen ydin (SC) |
|
||
| Lateraalinen | Lateraalinen preoptinen ydin | ||
| Lateraalinen ydin (LT) |
|
||
| osa supraoptisesta ytimestä (SO) |
|
||
| Tuberaalinen | Mediaalinen | Dorsomediaalinen hypotalamuksen ydin (DM) |
|
| Ventromediaalinen ydin (VM) |
|
||
Arcuate nucleus (AR)
|
|||
| Lateraali | Lateraalinen ydin (LT) |
|
|
| Lateraaliset tuberaaliset ytimet | |||
| Posteriorinen | Mediaalinen | Mammillaariset ytimet (osa mammillaarielimiä) (MB) |
|
| Takimmainen ydin (PN) |
|
||
| Lateraalinen | Lateraalinen ydin (LT) |
Ks. esim: Ventrolateraalinen preoptinen ydin
Lähdöt
Hypotalamuksen lähdöt voidaan jakaa kahteen kategoriaan: neuraalisiin projektioihin ja hormonaalisiin hormoneihin.
Neuraaliset projektiot
Useimmat hypotalamuksen kuitujärjestelmät kulkevat kahteen suuntaan (kaksisuuntaiset).
- Projektiot hypotalamuksen kaudaalisille alueille kulkevat mediaalisen etuaivokimpan, mammillotegmentaalisen radan ja dorsaalisen pitkittäisen fasciculuksen kautta.
- Projektiot hypotalamuksen rostraalisille alueille kulkevat mammillototalamuksen radan, fornixin ja terminaalisen strian kautta.
Endokriiniset hormonit
Hypotalamus vaikuttaa endokriiniseen systeemiin ja säätelee emotionaalista käyttäytymistä, kuten vihaa ja seksuaalista toimintaa. Suurin osa hypotalamuksen tuottamista hormoneista jakautuu aivolisäkkeeseen hypofyysiportaalijärjestelmän kautta. Hypotalamus ylläpitää homeostaasia, johon kuuluu verenpaineen, sykkeen ja lämpötilan säätely.
Hypotalamuksen ensisijaiset hormonit ovat:
| Nimi | Muut nimet | Lyhenteet | Sijainti | Toiminta |
|---|---|---|---|---|
| Kortikotropiinia vapauttava hormoni | Kortikotropiinia vapauttava tekijä, kortikoliberiini | CRH, CRF | parvocellulaariset neuroendokriiniset neuronit paraventrikulaarisessa ytimessä | vasopressiinin kanssa, stimuloi aivolisäkkeen etuosaa erittämään ACTH:ta |
| Dopamiini | Prolaktiinia inhiboiva hormoni | DA, PIH | neuroendokriiniset neuronit arcuate nucleus | estävät prolaktiinin eritystä aivolisäkkeen etuosasta |
| Gonadotropiinia vapauttava hormoni | Luteinisoivaa hormonia vapauttava hormoni | GnRH, LHRH | neuroendokriiniset neuronit mediaalisessa preoptisessa ja kaarevassa ytimessä | stimuloi aivolisäkkeen etuosaa erittämään LH:ta ja FSH:ta |
| Kasvuhormonia vapauttava hormoni | Kasvuhormonia vapauttava tekijä, somatokriini | GHRH, GHRF, GRF | arcuate nucleus neuroendokriiniset neuronit | stimuloi aivolisäkkeen etuosaa erittämään kasvuhormonia |
| Melatoniini | suprachiasmaattinen ydin | |||
| Somatostatiini | kasvuhormoni-inhiboiva hormoni, Somatotropiinin vapautumista estävä tekijä | SS, GHIH, SRIF | neuroendokriiniset neuronit periventrikulaarisessa ytimessä | estävät kasvuhormonin eritystä aivolisäkkeen etupuolelta |
| Tyrotropiinia vapauttava hormoni | Tyrotropiinia vapauttava tekijä, Thyroliberiini, Protireliini | TRH, TRF | parvocellulaariset neuroendokriiniset neuronit paraventrikulaarisissa ja anteriorisissa hypotalamuksen ytimissä | stimuloi anteriorista aivolisäkettä erittämään TSH:ta |
Seuraavasti: Hypokretiini
Ruoan saannin kontrollointi
Hypotalamuksen ventromediaalisen ytimen äärimmäinen lateraaliosa vastaa ruoan saannin kontrolloinnista. Tämän alueen stimulaatio aiheuttaa lisääntynyttä ruoan saantia. Tämän alueen molemminpuolinen vaurio aiheuttaa ruoan saannin täydellisen loppumisen. Ytimen mediaalisilla osilla on ohjaava vaikutus lateraaliseen osaan. Ventromediaalisen ytimen mediaalisen osan bilateraalinen vaurio aiheuttaa hyperfagiaa ja eläimen lihomista. Ventromediaalisen ytimen lateraalisen osan myöhempi leesio samassa eläimessä saa aikaan ruoanoton täydellisen loppumisen.
Tähän säätelyyn liittyy erilaisia hypoteeseja:
- Lipostaattinen hypoteesi – tämän hypoteesin mukaan rasvakudos tuottaa humoraalisen signaalin, joka on oikeassa suhteessa rasvan määrään ja joka vaikuttaa hypotalamukseen vähentääkseen ruoanottoa ja lisätäkseen energiatuotantoa. On ollut ilmeistä, että hormoni leptiini vaikuttaa hypotalamukseen vähentääkseen ruoan saantia ja lisätäkseen energiantuottoa.
- Gutpeptidihypoteesi – ruoansulatuskanavan hormonien, kuten Grp:n, glukagonien, CCK:n ja muiden väitetään estävän ruoan saantia. Ruoan joutuminen ruoansulatuskanavaan laukaisee näiden hormonien vapautumisen, joka vaikuttaa aivoihin tuottaakseen kylläisyyden. Aivoissa on sekä CCK-A- että CCK-B-reseptoreita.
- Glukostaattinen hypoteesi – ventromediaalisissa ytimissä sijaitsevan kylläisyyskeskuksen toimintaa ohjaa todennäköisesti glukoosin käyttö neuroneissa. On postuloitu, että kun niiden glukoosin käyttö on vähäistä ja näin ollen kun niiden välinen valtimoverenkierron glukoosiero on pieni, neuronien aktiivisuus vähenee. Näissä olosuhteissa ravintokeskuksen aktiivisuus on hallitsematonta ja yksilö tuntee nälkää. Ruoan saanti lisääntyy nopeasti laskimonsisäisellä 2-deoksiglukoosin antamisella, jolloin glukoosin käyttö soluissa vähenee.
- Termostaattinen hypoteesi – tämän hypoteesin mukaan ruumiinlämpötilan lasku tietyn asetusarvon alapuolelle stimuloi ruokahalua, kun taas lämpötilan nousu asetusarvon yläpuolelle estää ruokahalua.
Sukupuolidimorfismi
Monet hypotalamuksen ytimet ovat sukupuolidimorfisia, eli sekä rakenteessa että toiminnassa on selviä eroja urosten ja naaraiden välillä.
Joitakin eroja on havaittavissa jopa karkeassa neuroanatomiassa: huomattavin on preoptisella alueella sijaitseva sukupuolidimorfinen ydin, jota on vain miehillä. Suurin osa eroista on kuitenkin hienovaraisia muutoksia tiettyjen hermosolujoukkojen kytkeytyvyydessä ja kemiallisessa herkkyydessä.
Näiden muutosten merkitys voidaan tunnistaa urosten ja naaraiden välisistä toiminnallisista eroista. Esimerkiksi kasvuhormonin eritysmalli on sukupuolidimorfinen, ja tämä on yksi syy siihen, miksi monissa lajeissa aikuiset urokset ovat paljon suurempia kuin naaraat.
Vastaukset munasarjojen steroideihin
Muut silmiinpistävät toiminnalliset dimorfismit ovat aikuisten käyttäytymisvasteissa munasarjojen steroideihin. Miehet ja naiset reagoivat eri tavoin munasarjarakkuloiden steroideihin osittain siksi, että hypotalamuksen estrogeenille herkkien neuronien ilmentyminen on sukupuolisesti dimorfista, eli estrogeenireseptorit ilmentyvät eri neuronijoukoissa.
Estrogeeni ja progesteroni voivat vaikuttaa geeniekspressioon tietyissä neuroneissa tai indusoida muutoksia solukalvopotentiaalissa ja kinaasiaktivaatiossa, mikä johtaa moninaisiin ei-genomisiin solutoimintoihin. Estrogeeni ja progesteroni sitoutuvat vastaaviin ydinhormonireseptoreihinsa, jotka siirtyvät solun tumaan ja ovat vuorovaikutuksessa DNA:n alueiden kanssa, jotka tunnetaan nimellä Hormonivaste-elementit (HRE), tai kiinnittyvät toisen transkriptiotekijän sitoutumiskohtaan. Estrogeenireseptorin (ER) on osoitettu transaktivoivan muita transkriptiotekijöitä tällä tavoin, vaikka geenin proksimaalisella promoottorialueella ei ole estrogeenivaste-elementtiä (ERE). ER:t ja progesteronireseptorit (PR:t) ovat yleensä geenien aktivaattoreita, joiden mRNA ja sitä seuraava proteiinisynteesi lisääntyvät hormonialtistuksen jälkeen.
Miesten ja naisten aivot eroavat toisistaan estrogeenireseptoreiden jakautumisessa, ja tämä ero on vastasyntyneiden steroidien altistumisen peruuttamaton seuraus. Estrogeenireseptoreita (ja progesteronireseptoreita) löytyy pääasiassa neuroneista anteriorisessa ja mediobasaalisessa hypotalamuksessa, erityisesti:
- preoptisella alueella (jossa sijaitsevat LHRH-neuronit)
- periventrikulaarisessa ytimessä (jossa sijaitsevat somatostatiinineuronit)
- ventromediaalisessa hypotalamuksessa (jolla on merkitystä seksuaaliselle käyttäytymiselle).
Gonaaliset steroidit rottien neonataalielämässä
Neonataalielämässä gonadiset steroidit vaikuttavat neuroendokriinisen hypotalamuksen kehitykseen. Ne määräävät esimerkiksi naaraiden kyvyn normaaliin lisääntymiskiertoon sekä urosten ja naaraiden kyvyn osoittaa sopivaa lisääntymiskäyttäytymistä aikuiselämässä.
- Jos naarasrotalle ruiskutetaan kerran testosteronia syntymän jälkeisen elämän ensimmäisinä päivinä (sukupuolisteroidivaikutuksen ”kriittisen jakson” aikana), hypotalamus muuttuu peruuttamattomasti maskuliiniseksi; aikuinen rotta ei kykene tuottamaan LH-aallon nousua vasteena estrogeenille (mikä on naaraille ominainen piirre), mutta pystyy osoittamaan urospuolista seksuaalista käyttäytymistä (kiipeilemään seksuaalisesti vastaanottavaisen naaraan päälle).
- Heti syntymän jälkeen kastroitu urosrotta sitä vastoin feminisoituu, ja aikuinen osoittaa naaraspuolista seksuaalista käyttäytymistä vasteena estrogeenille (seksuaalinen vastaanottavaisuus, lordoosi}.
Androgeenit kädellisillä
Arsaslapsilla androgeenien kehitysvaikutus ei ole yhtä selvä ja seuraukset eivät ole yhtä täydellisiä. Tyttöjen ”tomboyismi” saattaa heijastaa androgeenien vaikutuksia sikiön aivoihin, mutta monien mielestä kasvatussukupuoli ensimmäisten 2-3 vuoden aikana on tärkein sukupuoli-identiteettiä määrittävä tekijä, koska tässä vaiheessa joko estrogeenilla tai testosteronilla on pysyviä vaikutuksia joko naaraan tai uroksen aivoihin, ja ne vaikuttavat sekä heteroseksuaalisuuteen että homoseksuaalisuuteen.
Paradoksaalista on se, että testosteronin maskuliinisoivia vaikutuksia välittää estrogeeni. Aivoissa testosteroni aromatisoituu (estradioliksi), joka on tärkein aktiivinen hormoni kehitysvaikutusten kannalta. Ihmisen kivekset erittävät suuria määriä testosteronia noin viikolta 8 sikiöiässä 5-6 kuukauteen syntymän jälkeen (samanlainen perinataalinen testosteronin nousu on havaittu monilla lajeilla), ja tämä prosessi näyttää olevan miehen fenotyypin taustalla. Äidin verenkierrosta peräisin oleva estrogeeni on suhteellisen tehotonta, osittain siksi, että steroidia sitovien proteiinien pitoisuudet verenkierrossa ovat raskauden aikana korkeat.
Muut vaikutukset hypotalamuksen kehitykseen
Sukupuolisteroidit eivät ole ainoat tärkeät vaikutukset hypotalamuksen kehitykseen; erityisesti varhaiselämän esipuberteettikautinen stressi määrittää aikuisen hypotalamuksen kyvyn vastata akuuttiin stressitekijään. Toisin kuin gonadien steroidireseptorit, glukokortikoidireseptorit ovat hyvin laajalle levinneet kaikkialla aivoissa; paraventrikulaarisessa ytimessä ne välittävät CRF-synteesin ja -erityksen negatiivista takaisinkytkentäohjausta, mutta muualla niiden roolia ei tunneta hyvin.
Ikääntymisen vaikutukset hypotalamukseen
Tutkimukset naarashiirillä ovat osoittaneet, että sekä supraoptinen ydin (SON) että paraventrikulaarinen ydin (PVN) menettävät noin kolmanneksen IGF-1R-immunoreaktiivisista soluista normaalin ikääntymisen myötä. Myös vanhat kalorisesti rajoitetut (CR) hiiret menettivät enemmän IGF-1R:n ei-immunoreaktiivisia soluja säilyttäen samalla samanlaiset IGF-1R-immunoreaktiivisten solujen lukumäärät verrattuna vanhoihin Al-hiiriin. Näin ollen Vanhat-CR-hiiret osoittavat suurempaa IGF-1R-immunoreaktiivisten solujen prosenttiosuutta, mikä heijastaa lisääntynyttä hypotalamuksen herkkyyttä IGF-1:lle verrattuna normaalisti vanheneviin hiiriin.
Katso myös
- HPA-akseli
- Hypotalamuksen hypofyysinen järjestelmä
- Hypotalamuksen aivolisäke-lisämunuaisjärjestelmä
- Hypotalamuksen vauriot
- Mediaalinen etuaivokimppu
- Neuroendokrinologia
- Preoptinen alue
Lisäkuvat
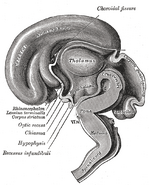
Lisää kuva tähän galleriaan
- http://www.cancer.gov/Templates/db_alpha.aspx?CdrID=46359
- http://www.sci.uidaho.edu/med532/hypothal.htm
- Kaavio ytimistä (psykedukaatio.org)
- Diagram of Nuclei (universe-review.ca)
- Diagram of Nuclei (utdallas.edu)
- Ellei taulukossa ole toisin mainittu, niin ref on: Guyton Eight Edition
- Walter F., PhD. Boron. Lääketieteellinen fysiologia: A Cellular And Molecular Approaoch, Elsevier/Saunders. Sivu 840
- http://thalamus.wustl.edu/course/hypoANS.html
- http://www.vivo.colostate.edu/hbooks/pathphys/endocrine/hypopit/overview.html
- Theologides A (1976). Anoreksiaa tuottavat välituotemetaboliitit. Am J Clin Nutr 29 (5): 552-8.
- John Money, ”The concept of gender identity disorder in childhood and adolescence after 39 years”, Journal of Sex and Marital Therapy 20 (1994): 163-77.
- Romeo, Russell D, Rudy Bellani, Ilia N. Karatsoreos, Nara Chhua, Mary Vernov, Cheryl D. Conrad ja Bruce S. McEwen (2005). Stressihistoria ja puberteettikehitys vuorovaikutuksessa muokkaavat hypotalamus-aivolisäke-lisämunuaisakselin plastisuutta. Endocrinology 147 (4): 1664-1674.
- Saeed O,Yaghmaie F,Garan SA,Gouw AM,Voelker MA,Sternberg H, Timiras PS. (2007). Insuliinin kaltaisen kasvutekijä-1-reseptorin immunoreaktiiviset solut säilyvät selektiivisesti kalorisesti rajoitettujen hiirten paraventrikulaarisessa hypotalamuksessa. Int J Dev Neurosci 25 (1): 23-8.
- Yaghmaie F, Saeed O, Garan SA, Voelker MA, Gouw AM, Freitag W, Sternberg H, Timiras PS (2006). Insuliinin kaltaisen kasvutekijä-1-reseptorin immunoreaktiivisten solujen ikäriippuvainen menetys supraoptisessa hypotalamuksessa vähenee kalorisesti rajoitetuilla hiirillä. Int J Dev Neurosci 24 (7): 431-6.
- F. Yaghmaie, O. Saeed, S.A. Garan, A.M. Gouw, P. Jafar, J. Kaur, S. Nijjar, P.S. Timiras, H. Sternberg, M.A. Voelker (2007). Hypotalamuksen IGF-1-herkkyyden muutosten seuranta ikääntymisen ja kalorimäärän rajoittamisen myötä. Experimental Gerontology 42 (1-2): 148-149.
Viitteet & Kirjallisuusluettelo
Artikkelit
- Arees, E.A. ja Mayer, B, (1967) Anatomical connections between medial and lateral regions of the hypothalamus concerned with food intake (Anatomiset yhteydet hypotalamuksen mediaalisten ja lateraalisten alueiden välillä koskien ruoan saantia), Science 157: 1574-5
- Grossman, S.P. (1960) Eating or drinking elicited by direct adrenergic or cholinergic stimulation of the hypothalamus, Science 132: 301-2.
- Hetherington, A.W. and Ranson, S.W. (1942) The relation of various hypothalamic lesions to adiposity in the rot, 76: 475-99.
Lisämateriaali
Artikkelit
- Google Scholar
.
- BrainMaps at UCDavis Hypothalamus
- Endokriininen järjestelmä ja hypotalamus
- High…Resolution Cytoarchitectural Primate Brain Atlases
- The Hypothalamus and Pituitary at endotexts.org
- Diagrammi ytimestä (psycheducation.org)
- Diagrammi ytimestä (universe-review.ca)
- Diagrammi ytimestä (utdallas.edu)
|
v-d-e
Ihmisen aivot: välikehä (TA A14.1.08, GA 9.807) |
||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Epithalamus |
|
Pinta
|
||||||||||||||
| Thalamus |
|
Pinta
|
Harmaa aine/
ytimet |
|||||||||||||
| Hypothalamus |
|
|||||||||||||||
| Subthalamus |
Subthalamic nucleus – Zona incerta |
|||||||||||||||
| Ventrikulaarinen järjestelmä: Kolmas kammio |
recesses: (Optinen syvennys, Infundibulaarinen syvennys, Suprapineaalinen syvennys, Pineaalinen syvennys) Hypotalamuksen sulcus – Kolmannen kammion Tela chorioidea Subfornikaalinen elin Apertuurat: Interventrikulaarinen/Monro Posterior commissure |
|||||||||||||||
|
v-d-e
Ihmisen anatomia, hormonitoiminta: Endokriiniset rauhaset |
|||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hypotalamuksen/ aivolisäkkeen akselit |
|
|
|
|
|||||||||||
| Pinealrauhanen |
Pinealosyytti – Corpora arenacea |
||||||||||||||
| Isletit. haima |
Alfasolu – Beetasolu – Deltasolu – PP-solu – Epsilon-solu |
||||||||||||||
ihmisen aivot: Limbinen järjestelmä
Amygdala – Cingulaarinen gyrus – Fornulaarinen gyrus – Hippokampus – Hypotalamus – Mammillaarirunko – Nucleus accumbens – Orbitofrontaalinen aivokuori – Parahippokampuksen gyrus