- Introduction
- The Genetic Basis of Hybrid Breakdown in Rice
- Molecular Mechanisms Underlying Hybrid Breakdown
- Hybridien hajoaminen tavanomaisessa riisinjalostuksessa
- Hybridien hajoaminen genomiikkaan perustuvassa jalostuksessa
- Keskustelu
- Data Availability Statement
- Author Contributions
- Interressiristiriita
- Kiitokset
Introduction
Kasvinjalostajat ja tutkijat ovat olleet kiinnostuneita hybridisaatiosta ja introgressiosta toisistaan poikkeavien genotyyppien välillä potentiaalisena ajurina, joka johtaa jälkeläisten ekologiseen eroavaisuuteen (Rieseberg et al., 2007; Arnold et al., 2012). Sitä vastoin hybridisaatioon voi joskus liittyä hybridikuntoisuuden heikkenemistä, kuten heikkoutta ja/tai steriiliyttä F1-sukupolvessa ja myöhemmissä sukupolvissa, jopa silloin, kun kyseessä on saman lajin jäsenten välinen hybridisaatio.
Vähentynyttä hybridin elinkelpoisuutta ja/tai hedelmällisyyttä, joka segregoituu F2-sukupolvessa tai myöhemmissä sukupolvissa, kutsutaan hybridin hajoamiseksi (hybrid breakdown, HB), johon liittyy väistämättä resessiivisiä alleeleja. Tämä lisääntymiseste on havaittu jo pitkään sekä kasveilla että eläimillä (Dobzhansky, 1970; Grant, 1971). Monissa tätä estettä koskevissa geneettisissä analyyseissä on käynyt ilmi, että se saadaan aikaan täydentävällä vaikutuksella lokusten välillä ja/tai lokusten välillä, joissa on erilaistuneita alleeleja, jota kutsutaan yleisesti Bateson-Dobzhansky-Muller (BDM) -inkompatibiliteetiksi (Rieseberg ja Willis, 2007). HB:hen liittyy välttämättä sisäisiä postzygoottisia lisääntymisesteitä, kuten hybridin elinkyvyttömyys (mukaan lukien heikkous, nekroosi ja kloroosi) ja hybridisteriliteetti (uroksen, naaraan tai molempien sukusolujen kohdalla); sen vuoksi jotkut tutkijat eivät välttämättä erota HB:tä F1-jälkeläisten elinkyvyttömyydestä ja steriiliydestä. Uskon kuitenkin, että HB on kätevä luokitus lisääntymisesteille, koska se viittaa niiden taustalla olevaan geneettiseen perustaan (eli resessiivisten alleelien osallisuuteen).
Uudemmat tutkimukset, joissa on käytetty Arabidopsis-mallikasvilajia, ovat antaneet paremman käsityksen HB:stä sen molekyylimekanismien osalta (Vaid ja Laitinen, 2019). Sitä vastoin, vaikka riisi (Oryza-laji) on mallikasvi, tämänhetkinen ymmärrys HB:n geneettisestä perustasta on edelleen rajallinen tässä lajissa, luultavasti siksi, että HB ei ole lisääntymiseste F1-hybrideissä, joissa odotetaan suurempaa jyväntuotantoa kuin sisäsiitosvanhempien linjoissa, ja luultavasti siksi, että F2- ja myöhemmissä sukupolvissa segregaatioon joutuvat huonommat kasvit voidaan helposti valikoitua fenotyypin perusteella pois jalostuspopulaatiosta. Myös resessiivisen periytymisen aiheuttama geneettisen kartoituksen vaikeus voi olla tämän rajoituksen taustalla.
Tässä hahmotan HB:tä riisin risteytyksissä viitaten Arabidopsis-tutkimusten antamiin tietoihin ja keskustelen siitä, miten HB:tä käsitellään riisinjalostuksessa.
The Genetic Basis of Hybrid Breakdown in Rice
On katsottu, että viljelty riisi, O. sativa japonica ja indica, muodostaa lajikompleksin niiden oletetun kantaisän (O. rufipogon) ja luonnonvaraisten lajien kanssa; niiden välisissä risteytyksissä on kuitenkin havaittu useita lisääntymisesteitä, mukaan lukien HB, (Oka, 1988; Vaughan ym,
Tietääkseni tohtori Oka oli ensimmäinen, joka kuvasi HB:n geneettisen perustan japonican ja indican välisissä intersubspesifisissä risteytyksissä, ja se selittyi kahdella komplementaarisella geenillä (Oka, 1957). Vastuullisia geenejä ei kuitenkaan kartoitettu kromosomeille, koska molekyylimarkkereita ei ollut tuolloin saatavilla.
Riisihybrideissä HB on usein kuvattu intersubspesifisissä (O. sativa ssp. japonica × ssp. indica) ja interspesifisissä (O. sativa × O. nivara ja O. sativa × O. glumaepatula) risteytyksissä (Oka, 1957; Sato ja Morishima, 1988; Wu ym., 1988), 1995; Li et al., 1997; Fukuoka et al., 1998, 2005; Kubo ja Yoshimura, 2002, 2005; Matsubara et al., 2007a,b, 2015; Yamamoto et al., 2007; Ichitani et al., 2012; intersubspesifisten risteytysten osalta; Sobrizal et al., 2001; Miura et al., 2008, interspesifisten risteytysten osalta) (taulukko 1). Suurin osa näistä HB-tapauksista johtui kahden lokuksen BDM-yhteensopimattomuudesta ja resessiivisistä alleeleista, jolloin 1/16 F2-jälkeläisistä, jotka olivat homotsygootteja resessiivisten alleelien suhteen molemmissa lokuksissa, osoitti HB-fenotyyppiä; 4/16 jälkeläisistä, jotka olivat heterotsygootteja vain toisessa lokuksessa, osoitti kuitenkin HB-fenotyyppiä risteytysyhdistelmästä riippuen (taulukko 1 ja kuva 1A). Takaisinristeytyshybrideissä yksi lokus oli jo kiinnittynyt toistuvan vanhemman alleeleihin; sen vuoksi 1/4 BCnF2-jälkeläisistä osoitti HB-fenotyyppiä (taulukko 1 ja kuva 1B).

Taulukko 1. BCnF2:n jälkeläiset. Riisin risteytyksissä raportoitujen hybridihäiriöiden geneettinen perusta.
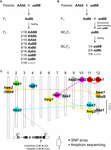
Kuva 1. Hybridin hajoamisen (HB) geneettinen perusta. (A) Kaavamainen esitys HB-genotyypistä, joka segregoituu F2-jälkeläisissä. Kun vanhempien genotyypit ovat AAbb ja aaBB, 1/16 F2-jälkeläisistä, joissa ei ole dominoivaa alleelia, joka on merkitty harmaalla merkillä (eli aabb), osoittaa alentunutta elinkelpoisuutta ja/tai hedelmällisyyttä, vaikka muut genotyypit ovat normaaleja. Joissakin riisin risteytyksissä F2-jälkeläisillä, joilla on vain yksi dominoiva alleeli (ts. Aabb, aaBb), on myös heikentynyt elinkelpoisuus ja/tai hedelmällisyys. (B) Kaavamainen esitys HB-genotyypistä, joka erottuu BCnF2-jälkeläisissä. Lisäksi 1/4 BCnF2-jälkeläisistä osoittaa alentunutta elinkelpoisuutta ja/tai hedelmällisyyttä. (C) HB:n taustalla olevien geenien kromosomaalinen sijainti riisiristeytyksissä. Geenit on karkeasti kartoitettu kunkin tutkimuksen geenikartoituksen tulosten perusteella. Joukko toisiaan täydentäviä geenejä on yhdistetty katkoviivalla. Komplementaarisia geenejä hbd1:n ja hwf1:n kanssa ei ole raportoitu. Esimerkki Yamamoto ym. (2010) julkaisemasta HB:tä aiheuttavasta funktionaalisesta nukleotidipolymorfismista on esitetty, ja tämän tiedon avulla voidaan tehdä tutkimus SNP-matriisilla tai amplikonisekvensoinnilla.
HB-fenotyyppi riisihybrideissä vaihtelee risteytysyhdistelmän mukaan. Esimerkiksi genotyyppi hbd2/hbd2 hbd3/hbd3 osoittaa heikkoutta, mutta ei selvää siemensteriliteettiä. Sen sijaan genotyypissä hbd4/hbd4 hbd5/hbd5 esiintyy sekä heikkoutta että siemensteriliteettiä (Matsubara ym., 2007a, 2015; Yamamoto ym., 2007; taulukko 1). On myös monimutkaisia tapauksia, joissa havaitaan sekä heikkoutta että siemensteriliteettiä osoittavaa HB:tä (genotyyppi hwe1/hwe1 hwe2/hwe2) tai vain siemensteriliteettiä osoittavaa HB:tä (hsa1/hsa1 hsa2/hsa2 hsa3/hsa3) samasta risteytysyhdistelmästä eli Asominori × IR24 -risteytyksestä huolimatta (Kubo ja Yoshimura, 2002, 2005; taulukko 1). Koska näillä HB:lla on itsenäinen geneettinen perusta, ainakin 5/64:llä F2-jälkeläisistä olisi HB-fenotyyppi tässä risteytyksessä.
Tähän mennessä useita HB:stä vastuussa olevia lokuksia on kartoitettu tietyille genomialueille DNA-markkereiden avulla japonica- ja indica-riisilajikkeiden välisissä risteytyksissä (Chr:t 7 ja 10, Fukuoka ym, 1998; Chrs 1 ja 12, Wu ym., 1995; Kubo ja Yoshimura, 2002; Matsubara ym., 2015; Chrs 6 ja 11, Fukuoka ym., 2005; Chrs 8, 9 ja 12, Kubo ja Yoshimura, 2005; Chrs 2 ja 11, Matsubara ym., 2007a; Yamamoto ym., 2007, 2010; Chrs 11 ja 12, Ichitani ym., 2012; taulukko 1 ja kuva 1C). Näiden tutkimusten tulokset osoittivat, että riisin HB:n taustalla olevat lokukset ovat yhteisiä joissakin risteytyksissä, mutta eroavat toisissa risteytyksissä, mikä viittaa niiden monimuotoistumiseen riisin genomissa. Esimerkiksi joissakin tapauksissa O. nivara -lajin, joka on läheisessä sukulaisuussuhteessa incica-lajikkeisiin, kantama HB-alleeli jakaa saman lokuksen indica-lajikkeen kanssa, mutta O. glumaepatula -lajin, joka on läheisessä sukulaisuussuhteessa viljeltyihin lajikkeisiin, kantama HB-alleeli ei jaa lokusta minkään muun lajikkeen kanssa (taulukko 1 ja kuva 1C). On huomattava, että edellä kuvatut HB-tapaukset johtuvat joukosta geenejä, joilla on merkittäviä vaikutuksia; on kuitenkin myös tapauksia, jotka johtuvat joukosta geenejä, joilla on vähäisiä vaikutuksia, esim. hieman heikentynyt siementen hedelmällisyys, joita ei voida jättää huomiotta riisinjalostuksessa, mutta joita ei ole vielä havaittu.
Molecular Mechanisms Underlying Hybrid Breakdown
Autoimmuunivaste: Bomblies et al. (2007) raportoivat ensimmäisenä, että autoimmuunivaste, joka NB-LRR-tautiresistenssigeeni tai R-geeni liittyy Arabidopsis HB:hen lajinsisäisissä risteytyksissä, vaikka tämä autoimmuunivaste ilmenee pääasiassa nekroosina F1-kasveissa, ja HB F2-jälkeläisissä näytti olevan lämpötilan ehdollistama. Arabidopsis-hybrideissä on kuvattu useita todisteita autoimmuunivasteen aiheuttamasta HB:stä (Alcázar ym., 2009, 2010). NB-LRR-geenien osallistuminen HB:hen viittaa siihen, että Arabidopsiksen HB:hen voi liittyä useita genomialueita; itse asiassa Chae ym. (2014) tekemä laaja tutkimus tukee tätä ajatusta.
Seuraavasti riisihybrideissä on raportoitu autoimmuunivasteen aiheuttamaa HB:tä japonica × indica -risteymässä (Yamamoto ym., 2010). Tässä tapauksessa HB esiintyy F2-kasveissa, kun indica-lajikkeen kantama kaseiinikinaasi I:tä koodaava hybrid breakdown 2 -geeni (hbd2) yhdistyy japonica-lajikkeen kantamaan R-geeniryhmään. Koska nämä aiheuttavat alleelit käyttäytyvät todennäköisesti osittain resessiivisinä, geenituotteet voivat olla osallisena (Matsubara ym., 2007a; Yamamoto ym., 2007, 2010).
Duplikoituneiden geenien vastavuoroinen vaimentaminen: Tätä käsitettä ehdotettiin ensimmäisen kerran teoreettisesti hybridien yhteensopimattomuuden geneettiseksi perustaksi (Werth ja Windham, 1991; Lynch ja Force, 2000). Mallissa yksi populaatio menettää toimintansa yhdessä lokuksessa ja säilyttää sen toisessa, kun taas toisessa populaatiossa vaikutus on päinvastainen. Näin ollen 1/16 F2-sygoteista ei ole toiminnallisia geenejä (Lynch ja Force, 2000). Bikard ym. (2009) ja Vlad ym. (2010) kuvaama HB Arabidopsis-hybrideissä on hyvä esimerkki tästä ilmiöstä. Tässä skenaariossa on raportoitu myös tapauksia, joihin liittyy epigeneettistä hiljentymistä (Agorio ym., 2017; Blevins ym., 2017).
Koska riisissä on esiintynyt sekä koko genomin että segmenttien monistumista (Wang ym., 2007; Guo ym., 2019), vastavuoroinen hiljentyminen voisi olla yleinen syy hybridiyhteensopimattomuuteen. Toistaiseksi ei kuitenkaan ole todisteita HB:stä, joka johtuisi duplikoituneiden geenien vastavuoroisesta hiljentämisestä riisissä.
Autoimmuunivasteesta ja vastavuoroisesta hiljentämisestä riippumatta on tunnistettu DEAD-box RNA-helikaasi 18:n ja toisen geenin (todennäköisenä ehdokkaana MORPHEUS MOLECULE 1) välinen vuorovaikutus, joka aiheuttaa HB:tä Arabidopsis-hybrideissä (Plötner ym., 2017; Vaid ym., 2020). Mielenkiintoista on, että HB-fenotyyppi vähenee F3- ja F4-sukupolvien välillä, mikä viittaa geeniekspression epigeneettiseen säätelyyn.
Riisihybrideissä kolmesta hybridi-steriliteetti-a (hsa) -lokuksesta Kubo et al. (2016) osoittivat hiljattain, että hsa1-lokus koostuu kahdesta geenistä ja että nämä geenit koodaavat DUF1618-proteiinia ja luonnostelematonta valkuaista, jolla on jonkin verran yhtäläisyyksiä nukleotidia sitovaan proteiiniin. Muiden komplementaaristen geenien, hsa2:n ja hsa3:n, molekyyliominaisuuksia ei ole raportoitu.
Hybridien hajoaminen tavanomaisessa riisinjalostuksessa
Vuosikymmenien ajan bulk-populaatiomenetelmää käytettiin laajalti tavanomaisessa riisinjalostuksessa. Tässä menetelmässä risteytyksen jälkeen F4- tai F5-populaatio kasvatetaan itsehedelmöittämällä irtotavarana (eikä yhden siemenen laskeutumismenetelmällä) ilman keinotekoista valintaa, mutta varhaisen sukupolven populaatio altistetaan luonnolliselle valinnalle ja elinkelpoisuusvalinnalle (Allard, 1960; Ikehashi ja Fujimaki, 1980 yksityiskohtaisesti). Bulkkipopulaatiomenetelmässä voidaan olettaa, että monet heikot ja/tai steriilit genotyypit poistuvat populaatiosta ennen varhaisemman sukupolven populaation perustamista; siksi tämä menetelmä voi antaa vain rajoitetusti tietoa HB:stä.
Riisinjalostajat ovat usein suorittaneet takaisinristeytyksen tai usean vanhemman risteytyksen ja sen jälkeen bulkkipopulaatiomenetelmän, kun haluttuja ominaisuuksia siirretään luovuttajalta/luovuttajilta lajikkeeseen. Empiirisesti on jo pitkään tiedetty, että näillä risteytysmenetelmillä voidaan lieventää lisääntymisesteiden aiheuttamaa valintakandidaattien häviämistä, koska nämä menetelmät usein vähentävät HB:hen liittyvien häiritsevien alleelikombinaatioiden erottumista hybridin jälkeläisissä. Lisääntymisesteiden lieventäminen näillä risteytysmenetelmillä on kuitenkin väistämättä riippuvainen vanhempien HB-genotyypistä.
Hybridien hajoaminen genomiikkaan perustuvassa jalostuksessa
Kasvien referenssikasvien genomisekvenssien julkaiseminen ja seuraavan sukupolven sekvensointitekniikoiden kehittyminen ovat kiihdyttäneet viljelykasvien molekyylijalostuksen etenemistä (Kole et al., 2015). Tässä yhteydessä viljelykasvien jalostuksessa on pohdittu genomiikka-avusteista jalostusta, kuten genominlaajuisten DNA-markkereiden genotyyppeihin perustuvaa genomivalintaa (Spindel ja Iwata, 2018).
Itsepölytteisten viljelykasvien genomiikka-avusteisessa jalostuksessa käytetään usein referenssipopulaatioina edistyneen sukupolven populaatioita, kuten rekombinanttisia sisäsiitoslinjoja, joista saadaan genominlaajuista genotyyppi ja fenotyyppitietoa. Tämän jälkeen varhaisen sukupolven jälkeläisissä (esim. F2) suoritetaan ainoastaan markkerigenotyyppeihin perustuva valinta. Tässä valintajärjestelmässä on huomattava, että vertailupopulaatio ei yleensä tarjoa tietoa HB:n genomialueesta, koska heikot ja/tai steriilit jälkeläiset olisi pitänyt eliminoida varhaisissa sukupolvissa risteytyksen jälkeen. Jos tällainen valintajärjestelmä otetaan käyttöön, joudumme siis luopumaan joistakin tärkeistä valintaehdokkaista. Vaihtoehtoisesti saatamme valita epätoivottuja ehdokkaita siten, että HB tulee näkyviin myöhemmissä sukupolvissa.
Keskustelu
Vaikka aihetta käsittelevien tutkimusten määrä on vähäinen, näyttää siltä, että HB:n perustana olevat lokukset riisin risteytyksissä ovat pikemminkin moninaisia kuin yhteisiä (taulukko 1 ja kuva 1). Kuten edellä on kuvattu, tietojen HB:hen liittyvien alleelien jakautumisesta risteytysvanhempien kesken pitäisi olla riisinjalostuksen edellytys, erityisesti genomiikka-avusteisen jalostuksen tehokkuuden maksimoimiseksi. Sen vuoksi tarvitaan lisää HB:n havaitsemista riisin risteytyksissä ja vastuullisten geenien kartoittamista riisin genomissa. Arabidopsis-hybrideissä tällaisia tietoja R-geeneistä ja niiden vuorovaikutuksessa olevista geeneistä on kartoitettu laajasti (Alcázar ym., 2010, 2014; Chae ym., 2014). Yamamoton ym. (2010) raportointi hbd2-geenistä riisihybridissä on hyvä esimerkki SNP-matriisien tai amplikonisekvensoinnin avulla tehdystä laajasta tutkimuksesta (kuva 1C). Myös hbd3- ja hsa1-geenit ovat ehdokkaita tämäntyyppiseen tutkimukseen. Lopulta sellaisten kriteerien kehittäminen, joiden avulla voimme ennustaa HB:n vanhempien linjojen genomitietojen perusteella, on tärkeä väline genomiikka-avusteisessa jalostuksessa. Vaikka kausaalista tekijää ei olisikaan tunnistettu yksittäisenä geeninä, läheisesti toisiinsa liittyvät markkerit, kuten yksinukleotidipolymorfismit, ovat tehokkaita HB:hen liittyvien haplotyyppien luokittelussa, koska viljellyn riisin linkitysepätasapainon on arvioitu ulottuvan 100-200 kb:iin, vaikka luonnonvaraisten lajien (kuten O. rufipogon) voi ulottua useiden kymmenien kb:n alueelle (Mather et al., 2007; McNally et al., 2009; Huang et al., 2010). Nämä tiedot mahdollistavat tehokkaamman ja vaikuttavamman risteytysyhdistelmän suunnittelun.
Bulked segreganttianalyysi, jota seuraa seuraavan sukupolven sekvensointi, voi olla hyödyllinen HB:hen liittyvien lokusten kartoituksessa sekä perinteisessä linkityskartoituksessa (kuten kvantitatiivisten ominaisuuksien lokusanalyysissä), koska HB-lajikkeen segreganteilla on usein erottuvia piirteitä (sellaisista piirteistä kuin kasvin korkeus, tillereiden lukumäärän määrä ja hedelmällisyys) kussakin risteytyspopulaatiossa olevista, tavanomaisesti kasvavista geeniperimästä. Kuvantamistietojen hankkiminen digitaalikameran ja drone-latauksen avulla voi myös olla tärkeässä asemassa fenotyypin määrityksessä laboratoriossa ja riisipellolla. Nämä toimet tarjoavat arvokasta tietoa paitsi riisinjalostajille myös evoluutiobiologeille.
Jos HB:hen liittyvä alleeli tunnistetaan, geenin häirintä ionisäteen tai genomin muokkauksen avulla voi lisäksi auttaa voittamaan HB:n, kuten F1-hybridien steriiliys riisissä on osoittanut (Koide ym., 2018; Xie ym, 2019).
Data Availability Statement
Kaikki tässä tutkimuksessa esitetyt tietokokonaisuudet sisältyvät artikkeliin/täydentävään aineistoon.
Author Contributions
Tekijä vahvistaa olevansa tämän työn ainoa tekijä ja on hyväksynyt sen julkaistavaksi.
Interressiristiriita
Tekijät ilmoittavat, että tutkimus on tehty ilman kaupallisia tai taloudellisia suhteita, jotka voitaisiin tulkita mahdolliseksi eturistiriidaksi.
Kiitokset
Omistan tämän artikkelin tohtori Darshan Brarin muistolle. Hän oli merkittävä riisinjalostaja, joka toi tarmokkaasti luonnonvaraisten lajien kantamia hyödyllisiä geenejä lajikkeisiin yli lisääntymisesteiden.
Allard, R. W. (1960). Principles of Plant Breeding. New York, NY: John Wily and Sons, Inc.
Google Scholar
Dobzhansky, T. (1970). Evoluutioprosessin genetiikka. New York, NY: Columbia University Press.
Google Scholar
Grant, V. (1971). Plant Speciation. New York, NY: Columbia University Press.
Google Scholar
Ikehashi, H., and Fujimaki, H. (1980). ”Modified bulk population method for rice breeding,” in Proceedings of the International Rice Research Conference on Innovative approaches to rice breeding, (Los Baños, PA: International Rice Research Institute), 163-182.
Google Scholar
Oka, H.-I. (1988). Viljellyn riisin alkuperä. Tokyo: JSSP.
Google Scholar